
 科普中国公众号
科普中国公众号
 科普中国微博
科普中国微博

 帮助
帮助
 中国科普博览
中国科普博览 
出品:科普中国
作者:吕泽龙(中国科学院动物研究所)
监制:中国科普博览
长颈鹿可能是动物园中的明星物种了,每个参观长颈鹿的人都会惊叹它们的长脖子。在我们普通人看来,所有的长颈鹿似乎都是一样的——长脖子,花斑纹。
但最近世界自然保护联盟(IUCN)发布了一份里程碑式的评估报告,正式确认:长颈鹿不止一个物种而是四个物种,包括北方长颈鹿,南方长颈鹿,网纹长颈鹿和马赛长颈鹿。其中一部分的区分依据就是它们身上的条纹颜色以及分布区域。比如北方长颈鹿和网纹长颈鹿就分布于非洲偏中部和东北部。

苏州上方山动物园的网纹长颈鹿
(图片来源:作者拍摄)

南方长颈鹿,如果仔细对比,我们会发现它们二者的花纹是有区别的
(图片来源:作者拍摄于天津动物园)
当然,要IUCN承认长颈鹿分为四个物种,并不只是依靠长颈鹿身上的斑纹不一样这一个证据这么简单。事实上,现代科学中要区分物种还挺复杂的——虽然我们从小在生物学中就了解了生殖隔离,但生殖隔离并不是区分物种的金标准。
一、为什么不能仅根据生殖隔离划分物种?
在最初的分类实践中,生物学家普遍把“能不能生出可育后代”当作物种分类的标准,这就是课本里常见的“生物学物种概念”(BSC,Biological Species Concept)。按照这个思路,两个群体如果交配后代大多不育,就说明它们之间存在生殖隔离(不同群体难以稳定产生可育后代)。
比如我们熟知的和驴子生下来的马骡和驴骡(前者是父亲为驴,后者是父亲为马,最常说的骡子是马骡,),就是几乎没有生殖能力的,即使偶有生殖,但从群体来看也是并不稳定的。或者是杂交物种的某一个性别不具备生殖能力,比如雄狮和雌虎生下来的狮虎兽则是雄性能力不会生,但是雌性则有重新和雄狮、雄虎生娃的案例。还有黄牛和家牦牛生下来的犏牛,也是雄性不能生,雌性可育继续生育。这些经典例子让BSC显得既直观又好用。

狮狮虎兽(雄狮和雌狮虎兽所生)
(图片来源:Wikipedia)
但是问题在于,大自然并没有这么简单。科学家就发现了许多不同物种生下来的后代依旧能繁殖的例子。比如在北极圈附近,就有棕熊和北极熊杂交然后生下来的“棕白熊”,这些熊依旧有生殖能力。在海豚科下面,一些不同物种的海豚之间甚至可以形成稳定的杂交群体。此外,北美洲的郊狼和灰狼也时常诞下具有生殖能力的后代。因此,如果我们教条地按照能不能“生出可育后代”这个标准划分物种的话,我们就会把许多原本各自独立演化的谱系合并到一起了。

棕熊和北极熊混血个体
(图片来源: TreeHugger)
因此,现代分类学对物种的分类采取了非常谨慎的态度。不仅会考虑到是否存在生殖隔离,还会考虑形态特征、地理与生态分布等多种因素,只有当多个因素指向同一个结论时,我们才会明确物种的划分。
二、物种到底要如何划分?
在多种划分物种的依据中,形态学无疑是最直观的证据。科学家首先会看骨骼、牙齿、体色与花纹等是否在不同群体中稳定可辨,这要求在同一地区、不同个体之间都能反复识别,尽量排除环境造成的可塑性差异。比如印度犀与爪哇犀,就在头骨与门齿形态上有系统性区别。我们在前文提到的北极熊和棕熊虽然仅仅分化才几十万年,但它们却形成了不同的头骨形态、臼齿、毛色和耳朵大小等,这也是将它们分为两个物种的依据。
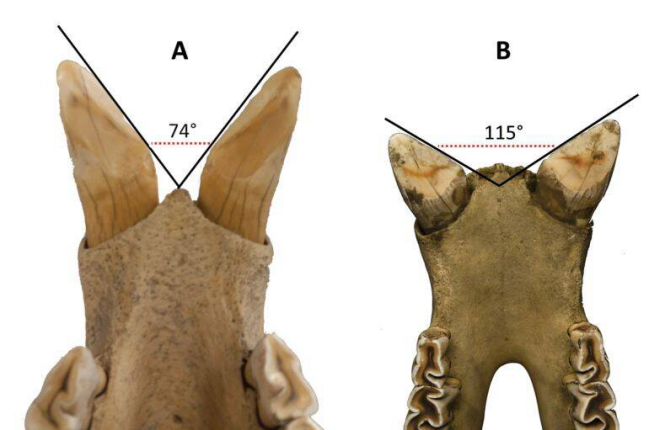
左侧为爪哇犀的门齿,右侧为印度犀
(图片来源:参考文献4)
遗传学就是深层证据之一。研究者会比较不同群体的DNA,看它们是否已经出现了长期的遗传分化并考察群体间的基因交换是否持久偏低。这里常用到核基因组和线粒体基因组的搭配:前者来自父母双方,能反映总体谱系关系;后者只随母系传递,能帮助识别历史上的雌系轨迹。以非洲大象为例,就被分为草原象与森林象,前者具有较大的耳朵和体重,不一样的头骨以及门齿形态,数量也比起后者多很多,分布区域也存在一定差别,但它们在核基因组上分化也非常显著,线粒体谱系也长期分开,说明两者不只是外形不同,而是在演化史上各走各路。遗传证据的意义在于,它能跨过短期的环境影响,给出时间尺度更长的分家账。

非洲草原象,拍摄于上海自然博物馆
(图片来源:作者)
当然,科学家也会考虑生殖层面的证据。但是现代研究更关心的是群体是否能长期维持独立,而不是单个杂交事件是否存在。虽然在前文提到的棕熊和北极熊、海豚、郊狼和灰狼等之间确实存在可育杂交的记录,但这些现象多出现在接壤狭窄地带或特定生态窗口,比例低,很难在大范围内把两个物种的基因库融合到一起。这些生殖隔离很模糊的不同物种,则会被统称为“物种复合体”,也称之为分类复合种(species complex)。它们形态学上有不少类似,亲缘上有些错综复杂,以至于难以确认种间界限。
当然,有时候两个物种之间的杂交种在长期演化之后,也会成功产生新的物种。比如俗称为狗熊的亚洲黑熊就是由熊科的两个不同支系混血杂交之后形成的新物种。还有黔金丝猴也是川金丝猴和其他金丝猴属支系混血而成,毛色继承了川金丝猴支系的黄色毛发也有其他金丝猴属的黑色毛。还有北美洲由郊狼和灰狼混血产生的赤狼(Canis rufus)这个物种(只分布在美国南部及其附近,极危物种)。

黔金丝猴,拍摄于北京动物园
(图片来源:作者)
最后还要看地理与生态的版图。不同群体如果长期分布在不同区域,被山脉、沙漠、洋流或气候带分隔,或者占据了不同的生态位,它们在行为、食性与微环境选择上就会逐渐拉开差距。这样的差距会反过来进一步降低基因交流,最后形成不同物种。在长颈鹿身上,我们看到不同谱系分布在非洲的不同地带,栖息偏好与活动范围并不相同,这些地理与生态差异与前述的形态和遗传证据一起,构成了同一个故事的多个侧面。
三、物种亚种的分化
但有时候,即使使用了这些综合证据,但不同证据之间还会存在不一致的情况,这是因为分化时间短或者生殖隔离不完全之类的原因,此时研究者就会使用“亚种”来记录分化。它与物种的区别在于:基因交流并未完全中断,遗传分化的深度较浅,外形差异虽然稳定,但不足以支撑“长期各自演化”的结论。多数亚种呈现地理分布上的分割,彼此之间常有过渡带,也就是在邻接区域能看到渐变或杂交个体)这说明它们仍属于同一物种的不同地域型。
例如我们熟悉的金钱豹就在IUCN上被列为8个亚种,包括非洲亚种(指名亚种),中国东北和华北,以及川陕甘等地的远东亚种,印度次大陆和斯里兰卡的亚种等。老虎也属于同一物种的多亚种格局:东北虎。华南虎,孟加拉虎等被认为起源于约7-11万年前的共同祖先,随后在气候变化与栖息地分化的背景下逐渐形成了各自稳定的头骨与被毛差异,这些差异在地域上能反复识别,但并未完全达到不同物种的门槛。

华南虎,毛色深一些且较短,拍摄于上海动物园
(图片来源:作者)

毛色浅一些且较为蓬松的东北虎,拍摄于长春动植物公园
(图片来源:作者)
但也有物种在讨论“要不要划亚种”的同时,发现内部已形成长期独立的谱系,但形态差异并不显著,因而尚未正式命名亚种。海洋中的大白鲨就是一个例子:基因组研究显示,其在十几万年前已分化为北太平洋、北大西洋和印度洋—太平洋三大谱系,这些谱系几乎互不通婚,但目前仍作为同一物种下的不同遗传支系来讨论,等待更多证据明确是否需要在命名层级上作进一步处理。
四、为什么要定好物种和亚种?
分类并不是书斋里的命名游戏,因为无论是研究还是保护,其资源都是有限的。把物种的边界厘清之后,哪一个物种更稀少,哪一个物种是关注重点,哪一些个体可以配对等问题就能更清晰了。更简单地来说,先认清楚不同物种的谱系,再避免外来血缘对本地物种的基因污染,这样保护和繁育才不会南辕北辙。
由物种不清导致的保护问题中的一个例子是全球只剩两只的一级保护动物斑鳖,这两只斑鳖均确认为雄性。斑鳖如此稀少,濒临灭绝,与它们在过去经常因为名字和分类地位,被和国家二级保护动物鼋搞混有关系。

苏州上方山动物园的斑鳖苏苏,全球仅剩下确认的斑鳖之一
(图片来源:作者)
回到文章开头的长颈鹿。这四种长颈鹿在分布区域、骨骼形态、核基因组和线粒体基因组等都存在较多的差别。且不同种类的长颈鹿的个体数量也不相同,其中分布在中非共和国,南苏丹等国家地区的北方长颈鹿数目只剩下7000多头,南方长颈鹿数目则超过6万头,而网纹长颈鹿和马赛长颈鹿在各自分布区也面对不同的生境破碎化与人为压力。现在,新的物种分类在更新后,配额、走廊建设、种群间调运与配对计划都会按物种分别制定,而不再用同样的标准来保护所有长颈鹿。这就能避免把资源过度分散导致的保护失调的问题。
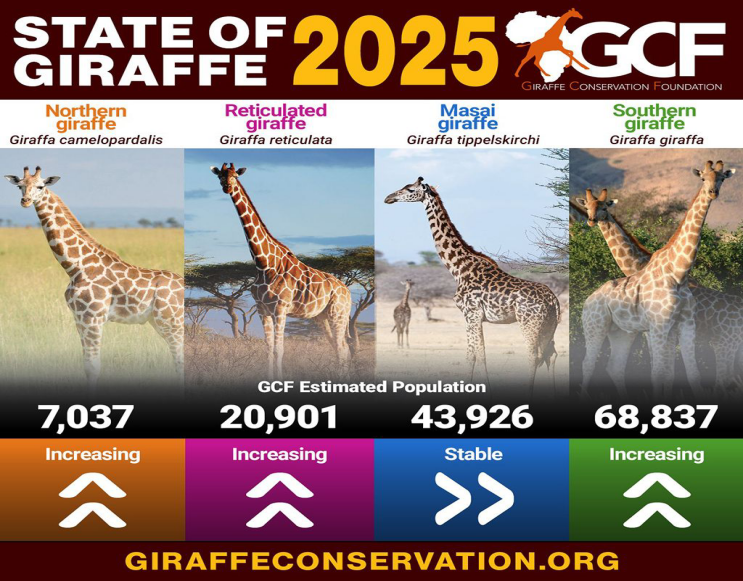
四种长颈鹿目前的数目,从左到右依次是北方长颈鹿,网纹长颈鹿,马赛长颈鹿和南方长颈鹿。其中马赛长颈鹿数目相对稳定,其他三种在增长
(图片来源:giraffeconservation)
参考文献:
1.Julian Fennessy et al. Multi-locus Analyses Reveal Four Giraffe Species Instead of One. Current Biology.2016; 26(18): P2543-2549.
2.Li HM, Liu BY, Shi MH, et al. Haplotype-resolved and chromosome-scale genomes provide insights into co-adaptation between the Amur tiger and Amur leopard. Zool Res. 2024;45(3):464-467.
3.Wang MS, Murray GGR, Mann D, et al. A polar bear paleogenome reveals extensive ancient gene flow from polar bears into brown bears. Nat Ecol Evol. 2022;6(7):936-944.
4.Nardelli F, Heißig K. A taxonomic review of the genus Rhinoceros with emphasis on the distinction of Eurhinoceros (Perissodactyla, Rhinocerotidae). Zookeys. 2025;1230:303-333.
5.Broggini C, Cavallini M, Vanetti I, Abell J, Binelli G, Lombardo G. From Caves to the Savannah, the Mitogenome History of Modern Lions (Panthera leo) and Their Ancestors. Int J Mol Sci. 2024;25(10):5193.
6.Amat F, Escoriza D. Biogeographic Inferences on the Evolutionary History of the King Cobra (Ophiophagus hannah, Cantor 1836) Species Complex. Zool Stud. 2022;60:e28
7.Wagner I, Smolina I, Koop MEL, et al. Genome analysis reveals three distinct lineages of the cosmopolitan white shark. Curr Biol. 2024;34(15):3582-3590.e4.
8.Das, Indraneil ; Gowri Shankar, P. ; Swamy, Priyanka et al. / Taxonomic revision of the king cobra Ophiophagus hannah (Cantor, 1836) species complex (Reptilia: Serpentes: Elapidae), with the description of two new species. In: European Journal of Taxonomy. 2024;961:pp.1.
9.Wu H, Wang Z, Zhang Y, et al. Hybrid origin of a primate, the gray snub-nosed monkey. Science. 2023;380(6648):eabl4997.
10.Zou T, Kuang W, Yin T, et al. Uncovering the enigmatic evolution of bears in greater depth: The hybrid origin of the Asiatic black bear. Proc Natl Acad Sci U S A. 2022;119(31):e2120307119.
11.Wang MS, Wang S, Li Y, et al. Ancient Hybridization with an Unknown Population Facilitated High-Altitude Adaptation of Canids. Mol Biol Evol. 2020;37(9):2616-2629.
12.Low WY, Tearle R, Liu R, et al. Haplotype-resolved genomes provide insights into structural variation and gene content in Angus and Brahman cattle. Nat Commun. 2020;11(1):2071.
13.Wilkins JS. What is a species? Essences and generation. Theory Biosci. 2010;129(2-3):141-148.
14.Curry CJ, Davis BW, Bertola LD, White PA, Murphy WJ, Derr JN. Spatiotemporal Genetic Diversity of Lions Reveals the Influence of Habitat Fragmentation across Africa. Mol Biol Evol. 2021;38(1):48-57.
15.Bertola LD, Jongbloed H, van der Gaag KJ, et al. Phylogeographic Patterns in Africa and High Resolution Delineation of Genetic Clades in the Lion (Panthera leo). Sci Rep. 2016;6:30807.
16.Rong RH, Cai HD, Yang XQ, Wei J. Fertile mule in China and her unusual foal. J R Soc Med. 1985;78(10):821-825.
17.Gaughran SJ, Gray R, Ochoa A, et al. Whole-genome sequencing confirms multiple species of Galapagos giant tortoises. Evolution. 2025;79(2):296-308.
18.Alcaide M, Scordato E S,Price T D, et al. Genomic divergence in a ring species complex. Nature,2014, 511(7507):83-85.
19.Mondol S, Moltke I, Hart J, et al. New evidence for hybrid zones of forest and savanna elephants in Central and West Africa. Molecular ecology, 2015, 24(24): 6134-6147.
20.Du H, Yu J, Li Q, Zhang M. New Evidence of Tiger Subspecies Differentiation and Environmental Adaptation: Comparison of the Whole Genomes of the Amur Tiger and the South China Tiger. Animals (Basel). 2022;12(14):1817.
21.Liu YC, Sun X, Driscoll C, et al. Genome-Wide Evolutionary Analysis of Natural History and Adaptation in the World's Tigers. Curr Biol. 2018;28(23):3840-3849.e6.
22.Pilot, M. Disentangling the admixed trails of grey wolf evolution. Molecular Ecology.2021;30, 6509–6512.
23.Hai-Meng Li, Bo-Yang Liu, Min-Hui Shi, Le Zhang, Shang-Chen Yang, Sunil Kumar Sahu, Liang-Yu Cui, Shan-Lin Liu, Nicolas Dussex, Yue Ma, Dan Liu, Wei-Yao Kong, Hao-Rong Lu, Yue Zhao, Love Dalén, Huan Liu, Tian-Ming Lan, Guang-Shun Jiang, Yan-Chun Xu. Haplotype-resolved and chromosome-scale genomes provide insights into co-adaptation between the Amur tiger and Amur leopard. Zoological Research.2024;45(3): 464-467.
24.Chai J, Lu CQ, Yi MR, et al. Discovery of a wild, genetically pure Chinese giant salamander creates new conservation opportunities. Zool Res. 2022;43(3):469-480.
25.Yan F, Lü J, Zhang B, et al. The Chinese giant salamander exemplifies the hidden extinction of cryptic species. Curr Biol. 2018;28(10):R590-R592.
26.Nie WH, Wang JH, Su WT, et al. Species identification of crested gibbons (Nomascus) in captivity in China using karyotyping- and PCR-based approaches. Zool Res. 2018;39(5):356-363.
27.Roca AL. African elephant genetics: enigmas and anomalies. J Genet. 2019;98:83.
28.Roca AL, Ishida Y, Brandt AL, Benjamin NR, Zhao K, Georgiadis NJ. Elephant natural history: a genomic perspective. Annu Rev Anim Biosci. 2015;3:139-167.

来源: 中国科普博览
内容资源由项目单位提供
