
 科普中国公众号
科普中国公众号
 科普中国微博
科普中国微博

 帮助
帮助
 科普中国创作培育计划
科普中国创作培育计划 
清晨的窗台上,一盆绿萝正舒展着叶片,它的茎秆以肉眼难以察觉的速度调整着弧度,缓慢地朝向光照的方向弯曲生长;而墙角中被阴影笼罩的吊竹梅也没有放弃寻找光明:纤细的茎节开始加速生长,直到突破阴影的边缘才放缓脚步。
刚播下的生菜种子在土表不会停滞太久,感应到光照后便开始慢慢萌发,而番茄的种子却要在黑暗的环境下才能安心发芽;豆芽在黑暗中会快速伸长,一旦接触到光照,就会发生形态变化并长成豆苗;菠菜只有在白天的长度超过13小时才开花,而菊花开花则需要日长短于一定的时间……
萌发、生长、开花……我们在生活中,常常能观察到植物的各个发育阶段都能“感应”到光照,并且对光的“喜好”也不同。但植物并不像我们人类一样有视觉,那他们是如何感受到光照的呢?
其实对于植物来说,光照有两方面的作用。一是大家所熟悉的 “光合作用”:植物通过“光合色素”捕获光能,将二氧化碳和水转化成储存着能量的有机物,并释放出氧气。除了可以作“能量来源”,光照还可以作为“信号”,影响、调控植物生长发育的全过程。而植物正是靠着体内的四类“光受体”蛋白,实现了对不同波长光信号的“精准解读”。
“追光者”——向光素
向光素是一种“蓝光受体”,主要接收400~500 nm波长的光。其上具有发色团,是分子能“捕捉”光的关键部位。向光素位分布在细胞膜上,就像是一名尽职尽责的指挥官一样,负责根据环境中的蓝光信号来指挥细胞的功能:
当向光素捕捉到外界的蓝光后,结构会发生变化(自磷酸化),这个过程非常迅速,比我们眨眼还要快上10倍;
接下来,自磷酸化的向光素会影响“生长素”运输蛋白质在膜上的分布,导致背光侧生长素浓度明显高于向光侧;
生长素的主要作用是促进细胞伸长,这种分布不均会导致植物背光侧伸长更快,最终朝向光源的方向弯曲生长。
所以我们常常能发现,不管如何改变光源的方向,植物总是能追着光生长。研究表明,向光素不仅能影响植物的生长方向,还可以调控气孔开闭、叶片伸展等多项生理功能。
“双重身份”——光敏色素
光敏色素有两种存在形态:“红光”吸收型(Pr),发色团吸收波长为600-700 nm;“远红光”吸收型(Pfr),发色团吸收波长为700-750 nm。在红光/远红光照射下,两种形态能够发生可逆的转换:
黑暗条件下,光敏色素以Pr(失活态)的形态存在于细胞质;
当环境中有红光时,Pr转变为Pfr;
Pfr为活性形态,可以从细胞质进入细胞核当中;
进入细胞核后,Pfr调控(开启或关闭)相关基因的表达,如启动种子萌发、调控开花等;
遇到远红光时,则变回失活态Pr。
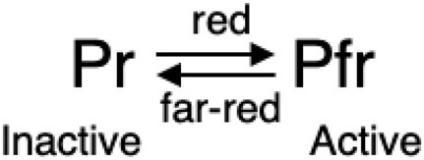
光敏色素两种形态的转换。图源:参考文献[3]
位于底层的植物幼苗正是通过检测环境中红光/远红光比例,判断上方是否存在“竞争对手”。当上方有其他植物时,红光被光合作用吸收,远红光占比增加,因此底层植物的Pfr水平降低,植物得以启动相关基因表达从而促进茎秆快速生长,争取早日接触到阳光。
“躲猫猫”高手——隐花色素
“隐花色素”这个名字的来源很有趣,早期人们最先在苔藓等隐花植物(不开花的植物)中发现了感知蓝光的能力,但是一直无法确定具体的受体是什么。经过一个多世纪的研究,科学家终于找到了这种不肯轻易露面的受体,因此起了“隐花色素”这个名字。
隐花色素的发色团能够接收近紫外光UV-A(320-400 nm)和蓝光(400-500 nm)。隐花色素主要有CRY1和CRY2两种亚型:CRY1在细胞质和细胞核中均有分布,CRY2主要存在于细胞核中。黑暗状态下,CRY2以单体的形式存在,蓝光照射后会迅速“抱团”形成同源二聚体,从而调控相关基因的表达。
隐花色素在植物中参与调控“形态建成”、“开花时间”等多项生理过程。比如即使在弱光环境中,隐花色素也可以敏锐捕捉蓝光,抑制下胚轴伸长,促进子叶扩展与叶绿体的形成,从而促进幼苗的生长发育。
隐花色素不仅仅存在于植物中,在我们人类当中,也有隐花色素的同源蛋白,与其他蛋白共同调节“昼夜节律”等生理过程。
“分子级防晒”——UVR8蛋白
UVR8蛋白是植物的紫外光受体,主要感知UV-B波段(280-315 nm)的光。与前3种受体不同的是,它没有专门的发色团,而是直接靠蛋白质上的色氨酸吸收光子。没有UV-B照射时,UVR8主要以同源二聚体的形式分布在细胞质中;受到UV-B照射后,二聚体解聚成有活性的单体,进入细胞核,调控相关基因的表达。UV-B能对植物造成DNA断裂等损伤,强度越大,核内UVR8单体积累越多,植物对紫外光的响应也越强烈。因此,UVR8可以一定程度上避免紫外线对植物造成过度损伤,堪称植物的“分子防晒霜”。同时,UVR8对植物的形态建成、侧根生长等也起到调控的作用。
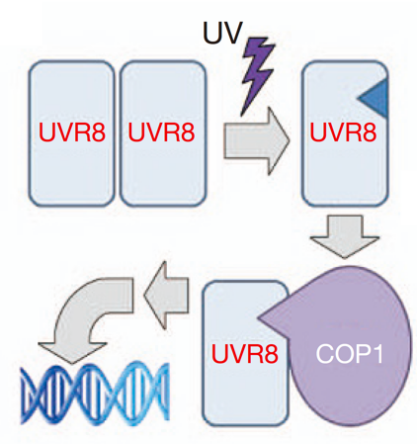
UVR8二聚体受到UV-B照射后解聚。图源:参考文献[5]
相信大家一定发现了,植物的感光机制基本遵循 “接收光信号→受体结构改变→调控基因表达→植物适应性提高”的过程。各种光受体之间相互配合,帮助植物精准感知环境中不同强度、不同波长的光线环境,共同调控植物的生长发育全过程。
利用植物的感光机制,也可以让我们的物质精神生活变得更加丰富多彩。比如适当补充蓝光来提高作物产量、通过改变光照时间让多种菊花统一开放迎接国庆佳节等等。
参考文献:
[1] Borthwick HA, Hendricks SB, Parker MW, Toole EH, Toole VK. A Reversible Photoreaction Controlling Seed Germination. Proc Natl Acad Sci U S A. 1952;38(8):662-666. doi:10.1073/pnas.38.8.662
[2] Kimura T, Tsuchida-Mayama T, Imai H, Okajima K, Ito K, Sakai T. Arabidopsis ROOT PHOTOTROPISM2 Is a Light-Dependent Dynamic Modulator of Phototropin1. Plant Cell. 2020;32(6):2004-2019. doi:10.1105/tpc.19.00926
[3] Huq E, Lin C, Quail PH. Light signaling in plants-a selective history. Plant Physiol. 2024;195(1):213-231. doi:10.1093/plphys/kiae110
[4] Wang Q, Zuo Z, Wang X, et al. Photoactivation and inactivation of Arabidopsis cryptochrome 2. Science. 2016;354(6310):343-347. doi:10.1126/science.aaf9030
[5] Wu D, Hu Q, Yan Z, et al. Structural basis of ultraviolet-B perception by UVR8. Nature. 2012;484(7393):214-219. Published 2012 Feb 29. doi:10.1038/nature10931
策划制作
作者:陈一欧 科普作者
审核:王康 北京植物园科普中心主任 北京植物学会副理事长 中国植物学会会员
来源: 科普中国创作培育计划
内容资源由项目单位提供
