图文
版权归原作者所有,如有侵权,请联系我们
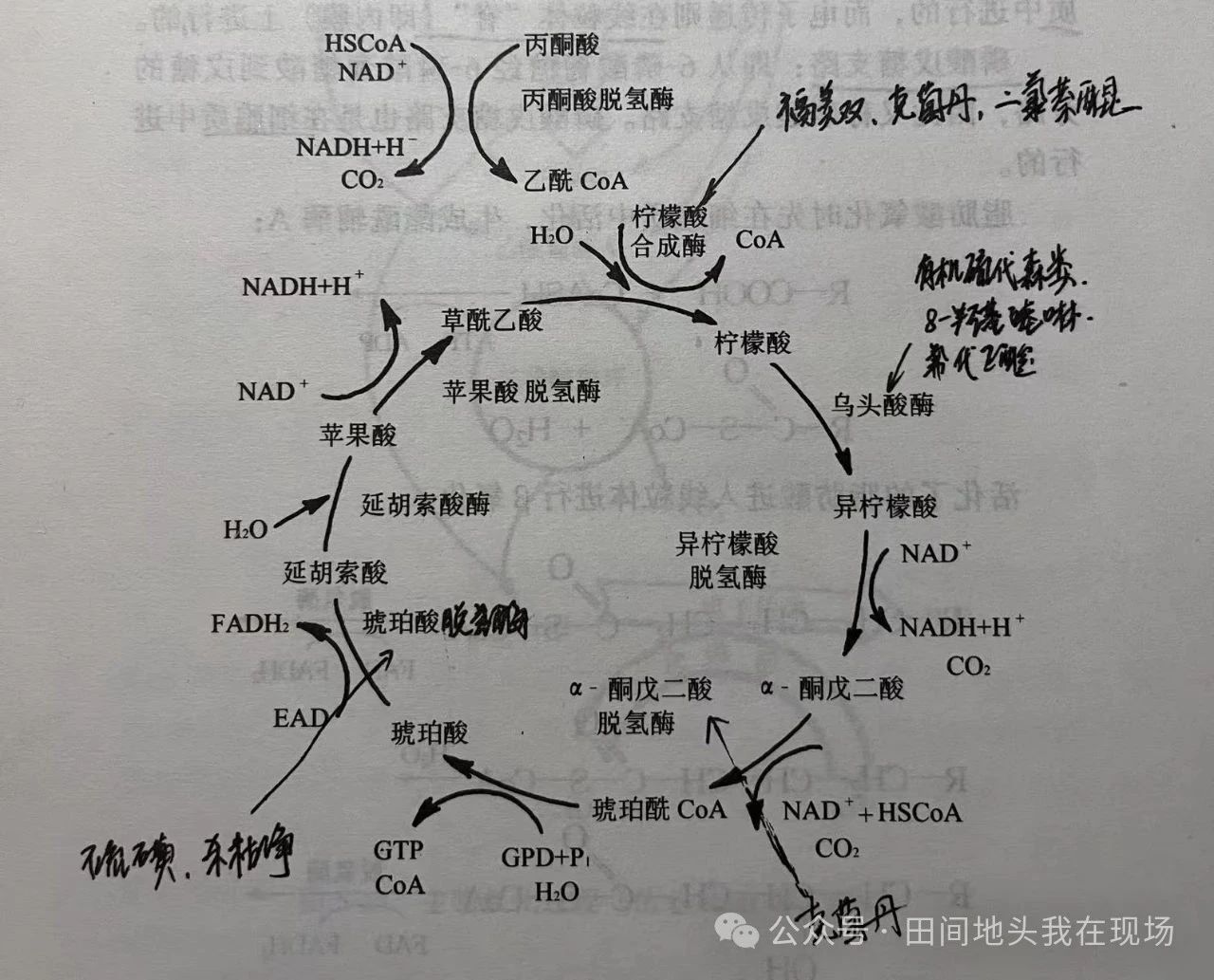
图文简介: 噻菌灵probenazole对稻瘟病和白叶枯病有良好的防治效果,噻菌灵和其在稻株中的主要代谢产物糖精(邻磺酰苯甲酰亚胺)和水杨酸,可以促使有毒酯类物质的产生并提高过氧化物酶的水平,阻止病菌扩展。
来源: 公众号:田间地头我在现场

 科普中国公众号
科普中国公众号
 科普中国微博
科普中国微博

 帮助
帮助
 一芳田自然科普
一芳田自然科普