
 科普中国公众号
科普中国公众号
 科普中国微博
科普中国微博

 帮助
帮助
 中国科普博览
中国科普博览 
出品:科普中国
作者:李煜,赵珺(中国科学院植物研究所)
审核:刘永秀(中国科学院植物研究所)
监制:中国科普博览
编者按:为解码生命科学最新奥秘,科普中国前沿科技项目推出“生命新知”系列文章,从独特的视角,解读生命现象,揭示生物奥秘。让我们深入生命世界,探索无限可能。
在我们的蓝色星球上,有着千姿百态的植物。春兰,夏荷,秋菊,冬梅……它们点缀了四季,也丰富了我们的生活。
种子,是植物智慧的一种体现,它们承载着种群的希望,通过多种方式传播到适宜生长的地方,并在恰当的时间萌发,从而实现植物种群的延续。种子的传播和萌发机制,展现了自然界中植物适应环境的奇妙智慧,也是科学界长期关注的重要课题。
种子的传播智慧花样百出
种子植物进化出了各种利于种子传播的方式,利用自然界中的各种力量来传播后代,让种子得以去往适宜萌发的环境和空间,如:风力传播、水流传播、弹射传播和动物传播等。
有些植物为种子制作了“小翅膀”,好风凭借力,送它去远方;有些种子则是天生的“游泳健将”,可以漂浮在水面上,随波逐流到新环境中生根发芽,如椰子和莲蓬等。

风力传播和水流传播
(图片来源:Veer图库)
动物传播可以看作是植物与动物的一场交易。动物吃掉植物的果实,然后在其它地方将种子排出。这样一来,动物获得了食物,种子则得到了“车票”。还例如,松鼠收集储藏松子,这些松子或被食用或被遗忘或被丢失。其中,从鼠口“逃生”的松子就可能在新的地方生根发芽。除了这些,采用动物传播方式的还有强行搭便车的苍耳子。

动物传播
(图片来源:参考文献1)
此外,有些植物的果实成熟后会爆裂,将种子向四方弹射。喷瓜、豌豆和油菜种子就是这种传播方式。
弹射传播(视频来源:Smithsonian Channel的图片转换)
在漫长的进化过程中,为了适应海岸潮间带潮起潮落的不稳定环境,红树林植物进化出了独特的“胎生”繁殖策略。这种策略使得种子在果实内部就开始萌发,发育成棒状的胚轴。当胚轴发育到一定程度后,它会脱离母树,在重力的作用下掉落到海滩的淤泥中,迅速扎根并长出新的个体。这种胎生现象是红树林植物对海岸生境的一种重要适应,它保证了植物在高盐、缺氧的沼泽地区能够有效地繁殖和生存。

红树林“胎生”植物
(图片来源:Veer图库)
种子休眠的形成与解除,大有智慧!
不仅动物会休眠(如鱼虾受精卵的休眠特性),植物界的种子也会休眠。
种子休眠是一种生物学特性,使得有活力的种子即便在适宜的环境条件下也不能萌发。这种特性是植物在长期进化过程中,为了抵御不利自然环境而形成的一种自我保护机制。种子休眠有助于延长种子的寿命,增加其传播距离,从而扩大植物种群的空间分布。此外,休眠状态还能防止种子在短时间内大量集中萌发,减少种内竞争,提高种群适应环境变化的能力。
在同一物种中,不同生态型的种子休眠也千差万别。以模式植物拟南芥为例,Col-0生态型的种子休眠期较短,大约只有2周左右,而Cvi生态型的种子则有较长的休眠期,可达半年之久。这种差异可能与Cvi生态型原产地的气候条件有关,它原生长于非洲的热带沙漠地区,那里的环境条件相对恶劣。种子休眠是一种关键的生物学策略,它保障种子能在合适的时机和环境中萌发,从而提高生存和繁衍的成功率,是植物适应环境的一种精巧的自我保护机制。

生态型不同导致种子休眠差异
(图片来源:杨月提供)
在种子的发育后期,伴随着种子的脱水过程,种子休眠逐渐形成。研究发现,种子形成过程期间的环境温度,对种子休眠的程度有显著影响。例如,模式植物拟南芥较低温度下形成的种子,休眠水平高;反之,如果温度高,种子休眠水平则低。这与低温诱导休眠因子DOG1积累有关,促进了种子休眠加深。
此外,植物激素对种子休眠的形成也有重要影响,它是植物体内产生的极其微量的化合物,能调节很多生理过程。其中,脱落酸在种子休眠形成中起关键作用,而赤霉素则在促进种子萌发中非常重要,二者的相对平衡决定了种子休眠和萌发。值得注意的是,赤霉素并不能促进所有植物种子的萌发。例如,寄生植物的种子就进化出了独特的机制,它们依赖于感受宿主分泌的独脚金内酯(strigolactone,SL)来启动萌发。独脚金内酯是一类源自类胡萝卜素的倍半萜类植物激素,它具有调控植物分枝,促进植物和丛枝菌根真菌共生的功能,因能够促进寄生植物独脚金种子的萌发而得名。
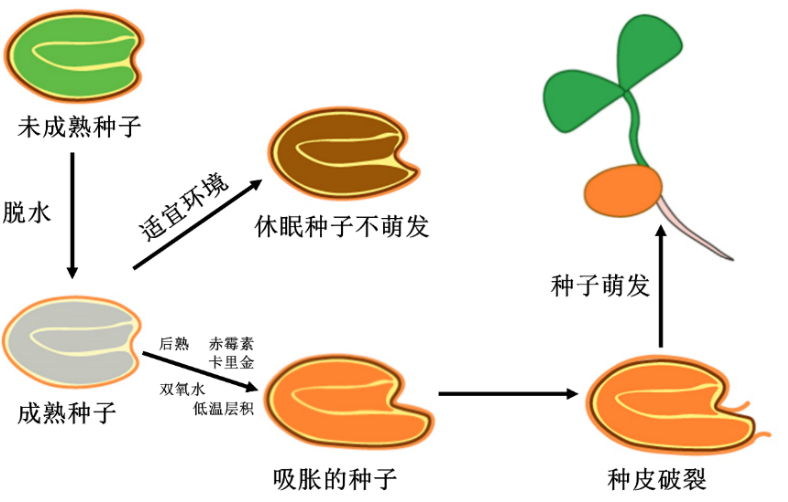
种子休眠和萌发
(图片来源:作者根据文献绘制)
种子携带着亲代植物生长发育过程中的记忆,其特殊的结构也如同设置了密码一般,在一定程度上阻碍了与水分、气体等萌发必要条件的接触,从而形成休眠状态。为了简化理解,我们可以将种子休眠主要分为两类:一类是种子物理性休眠,这种休眠是由种皮或果皮的结构特点导致的,它们影响种子的吸水、透气性能,或者限制了胚的生长;另一类是种子生理性休眠,这种休眠是由种子胚本身的原因所引起的,包括胚发育未完成、生理上未完成后熟,以及存在抑制萌发的物质。
在自然界中,种子解除休眠也是一个充满智慧的过程。种子会充分利用四季更迭、风霜雨雪、光温周期以及动物啃食和微生物作用等自然环境变化,来有效解除物理性休眠和生理性休眠,以待合适的萌发时机。
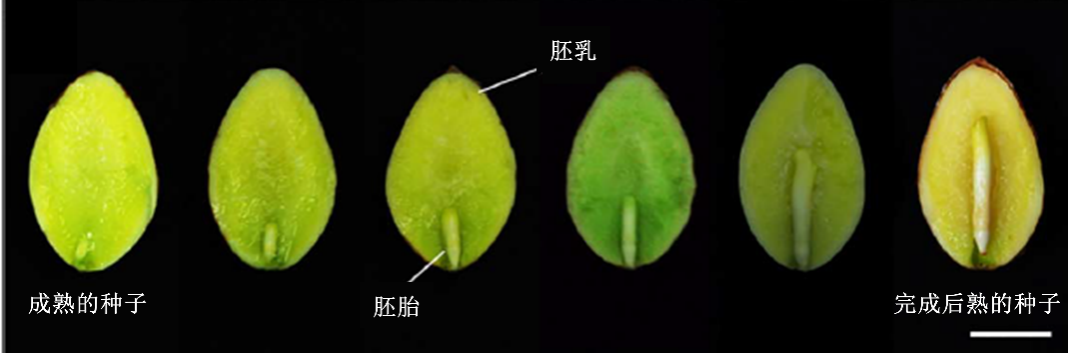
逐渐后熟的银杏种子
(图片来源:参考文献2)
渡渡鸟和大颅榄树的故事就是动物帮助植物种子萌发的一个经典例子。渡渡鸟,这种曾经生活在毛里求斯岛上的不会飞的鸟类,由于西方殖民者的到来,遭受了肆意捕杀,最终走向灭绝,继而引起了岛上生态环境的恶化。大颅榄树是毛里求斯岛上的一个树种,它的种子需要借助渡渡鸟的消化系统来削弱种(果)壳,以便顺利萌发。渡渡鸟的灭绝导致大颅榄树的种子无法得到适当的处理,继而也引发了大颅榄树丛林的逐渐凋亡。
直到1981年,科学家们发现,通过模仿渡渡鸟的消化过程,可以促进大颅榄树种子的萌发。这一发现为大颅榄树的保育工作带来了希望,也让人们意识到,生态系统中的每一环都是非常重要的,某一动植物的缺失都可能引起不可挽回的连锁反应。目前,世界上仅存的渡渡鸟软组织标本保存在牛津大学自然史博物馆,该博物馆还以渡渡鸟的卡通形象作为标志,以此来纪念这一物种,并提醒人们关注生物多样性的保护。
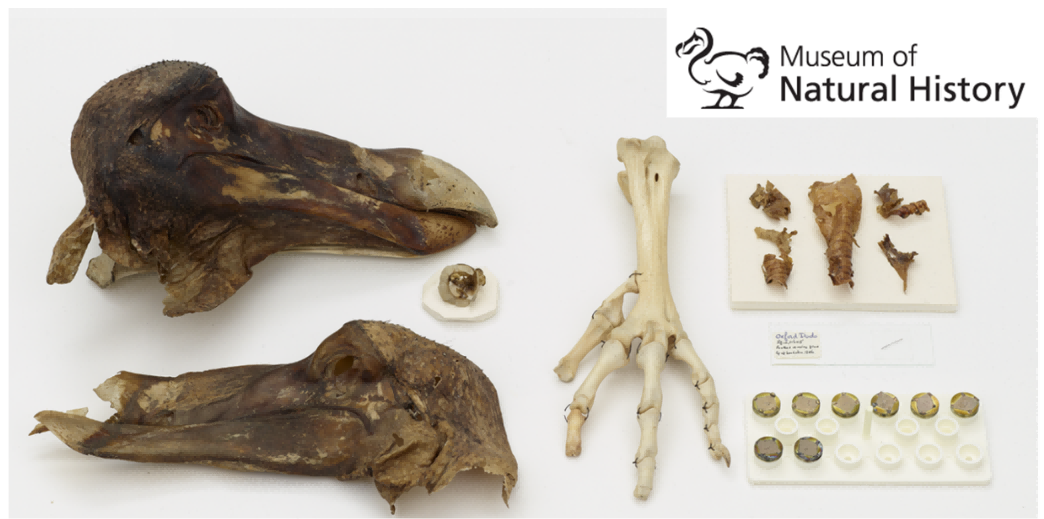
渡渡鸟软组织标本和牛津大学自然史博物馆标志
(图片来源:Museum of Nature History)
种子会通过感受光温,调控生命进程
安家落户的种子,静待休眠解除,开启生命的下一阶段。水分与适宜的温度是种子能够萌发的必要条件。温度不仅向种子提供了季节和局部微环境的信息,还影响着萌发进程中酶促反应的进行。种子通过感受温度的变化,相应不断调整着其生命进程。
在对模式植物拟南芥的研究中,科学家们发现了光敏色素phyB不仅是植物感知红光的受体,起着植物的“眼睛”的作用,还是感知温度的受体。当phyB功能丧失后,种子萌发对高温胁迫就变得更加敏感,还会导致种子萌发率急剧下降。原因在于,高温会使活性phyB的含量逐渐减少,并引起种子中萌发抑制物ABA含量的升高。
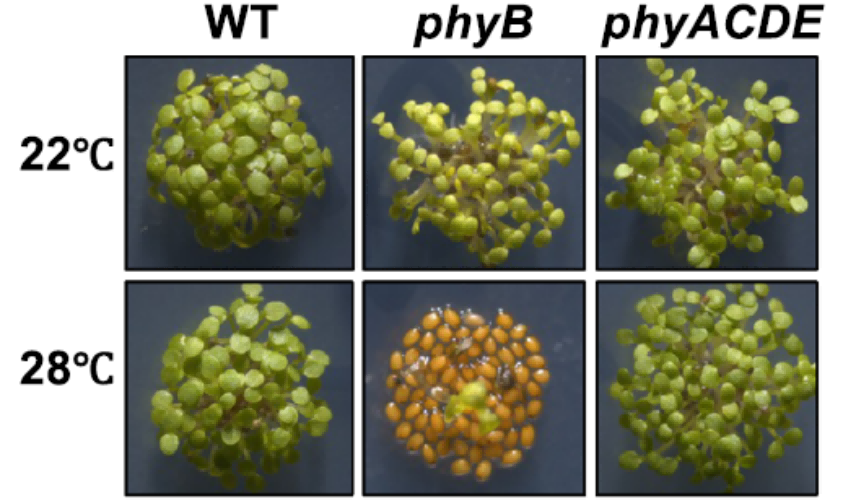
种子萌发对温度的敏感性
(图片来源:参考文献3)
野火烧不尽,春风吹又生:种子的分子智慧
森林火灾后,不需要人工干预,植被也会逐渐自我修复。植物在焚烧时,它们的身躯虽然化为了灰烬,但是这些灰烬中隐藏了新生命诞生的秘密。燃烧的植物会产生一种名为卡里金(Karrikin)的小分子化合物。随着大火后的第一场大雨,卡里金会渗入土壤中,强烈地促进土壤里种子的萌发。

火灾后森林的恢复情况
(图片来源:参考文献4)

森林灰烬中的卡里金促进种子萌发
(图片来源:参考文献5)
种子萌发后的生长也是充满巧思
种子萌发后,幼苗需要适应黑暗环境并克服土壤的机械压力才能破土而出。当它完全在土里时,幼苗依靠“油箱”中仅剩的营养“埋头”向上冲刺。子叶闭合向下弯曲的“埋头”状态,减少了背地生长遇到的土壤压力,还保护了脆弱的顶端分生组织。破土而出时,“埋头”状态还有利于顶开幼苗上方的重物,进一步保护幼苗。最终,幼苗打开子叶,开始土壤外的生活。“如果不落在肥土中而落在瓦砾中,有生命的种子决不会悲观,叹气,它相信有了阻力才有磨炼。”小小的种子居然深藏如此大智慧!

幼苗破土过程
(图片来源:参考文献6)
结语
经过亿万年的演化,稳定的、适合生存的生态系统逐渐形成。在这些生态系统中,每种生命形式都为了自身的生存和繁衍,进化出了独属自己的生存哲学,以躲避或抵御不良环境,影响适应环境,维持与环境的和谐。
种子植物,作为植物界中高度进化的种群,构成了地球表面绿色植被的主体。它们在种子的形成、结构、形状、传播方式以及种子休眠和萌发机制等方面,无不体现出大自然的巧思。
而这,只是大自然丰富生物资源的冰山一角,大自然还有更多的“智慧”有待我们发掘、学习、研究。未来,路还很长,探路者还在前行。
参考文献:
1. Falcón, W., Moll, D., and Hansen, D. M. (2020). Frugivory and seed dispersal by chelonians: a review and synthesis. Biological Reviews, 95, 142-166.
2. Jia, Z., Zhao, B., Liu, S., Lu, Z., Chang, B., Jiang, H., Cui, H., He, Q., Li, W., and Jin, B. (2021). Embryo transcriptome and miRNA analyses reveal the regulatory network of seed dormancy in Ginkgo biloba. Tree Physiology 41, 571-588.
3. Piskurewicz, U., Sentandreu, M., Iwasaki, M., Glauser, G., and Lopez-Molina, L. (2023). The Arabidopsis endosperm is a temperature-sensing tissue that implements seed thermoinhibition through phyB. Nature Communications 14, 1202.
4. Abella, S.R., and Fornwalt, P.J. (2015). Ten years of vegetation assembly after a North American mega fire. Global Change Biology 21, 789-802.
5. Chiwocha, S.D., Dixon, K.W., Flematti, G.R., Ghisalberti, E.L., Merritt, D.J., Nelson, D.C., Riseborough, J.-A.M., Smith, S.M., and Stevens, J.C. (2009). Karrikins: a new family of plant growth regulators in smoke. Plant Science 177, 252-256.
6. Shi, H., Lyu, M., Luo, Y., Liu, S., Li, Y., He, H., Wei, N., Deng, X.W., and Zhong, S. (2018). Genome-wide regulation of light-controlled seedling morphogenesis by three families of transcription factors. Proceedings of the National Academy of Sciences of the United States of America 115, 6482-6487.

来源: 中国科普博览
内容资源由项目单位提供
