
 科普中国公众号
科普中国公众号
 科普中国微博
科普中国微博

 帮助
帮助
 中国科普博览
中国科普博览 
出品:科普中国
作者:喜马拉雅小熊猫
监制:中国科普博览
常言道:“靠人不如凭己”。然而,在自然界中,有着这么一群植物,它们通过寄生,依靠吸收寄主的营养而存活发育。乍一听,是不是有点“寄生虫”那味儿了。今天,我们就一起揭开这些“靠自己不如靠别人”的植物们的“神秘面纱”。
什么是寄生植物
一般来说,两个营共生(Symbiotic relationship)生活的物种间有6种可能的关系,分别是互利共生(Mutualism)——双方都从共生关系中受利,比如地衣中的真菌和藻类;偏利共生(Commensalism)——共生关系对一方有利而对另一方无明显害处,比如鮣鱼和蝠鲼;偏害共生(Amensalism)——共生关系对一方有害而对另一方无明显利处,比如青霉与某些细菌;共栖(Neutralism)——共生关系对任何一方都无明显利害,比如偕老同穴与俪虾;竞争(Competition)——两者争夺共同的生活资源,对共生的双方都有害,比如水稻和稗;寄生(Parasitism)——寄生者寄附于另一种生物,利用寄主的养分生存,比如肉苁蓉和梭梭。
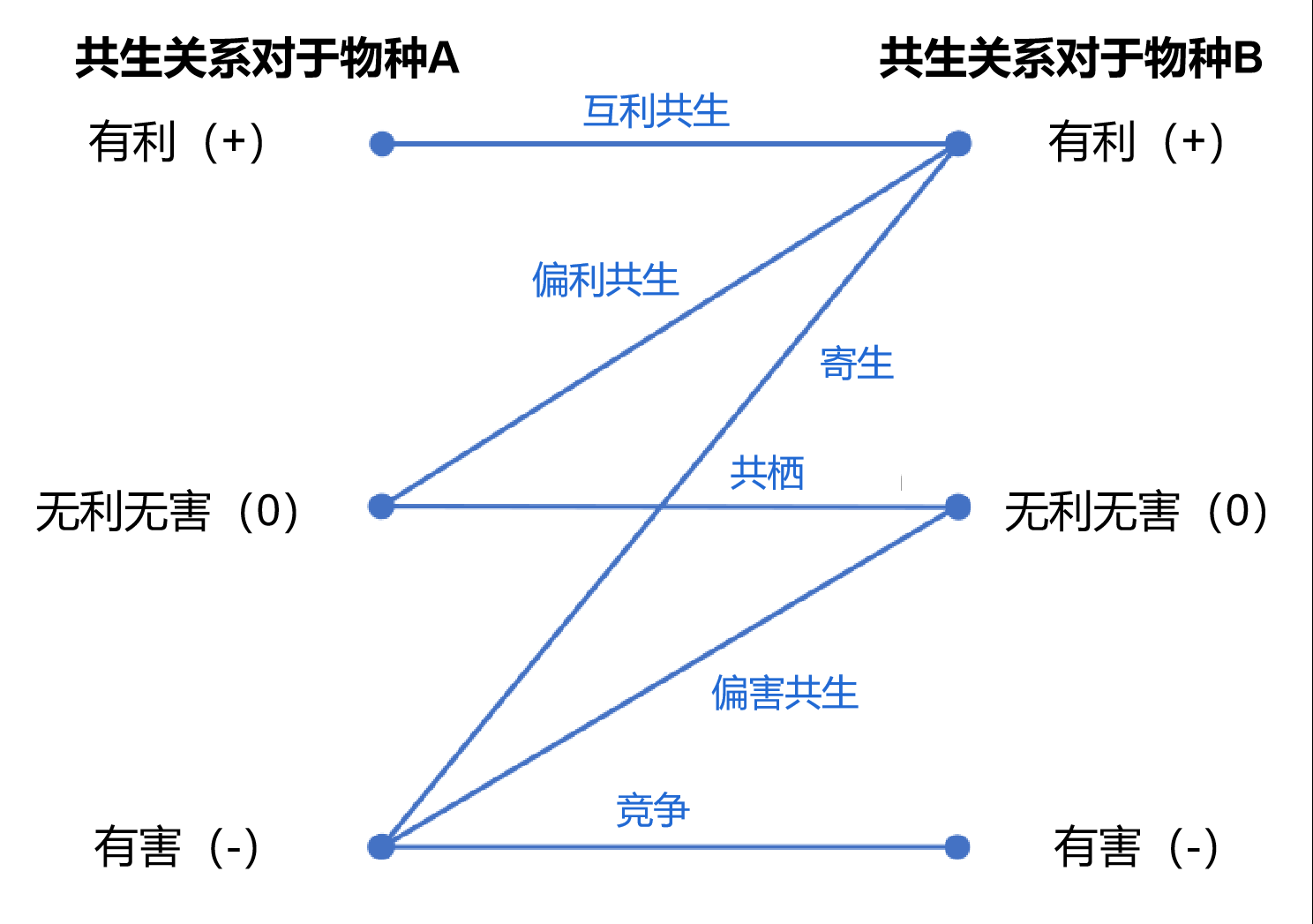
六种共生(Symbiosis)关系
(图片来源:https://en.wikipedia.org/wiki/Biological_interaction)
其中,寄生是最为独特的一种共生方式,这种共生方式在所有生物中广泛存在,包括原核生物、古菌、动物、植物、真菌以及其他真核生物。营寄生生活的生物通常会在演化过程中产生形态、生理、基因等各个方面的变化。
如今,我们已经认识到约292属和4750种寄生植物。在12个寄生植物类群中,檀香目是含有寄生植物最多的目,而列当科是含有寄生植物最多的科,它们共同向我们揭示了寄生植物的起源与演化过程。除此之外,在于寄主的共生过程中,寄生植物也与寄主进行着共同进化。
近年来,随着科学家们对于植物生理学及形态学的了解更加深入,通常认为广义的寄生植物包含两类,分别是吸器寄生植物和菌根异养植物。
虽然两者系统发育关系不同,但这两种类型的异养植物在生理学、解剖学、发育和生活史上具有许多相似性。
不同的是二者获取营养的方式,吸器寄生植物通常通过根与茎特化而成的吸器直接侵入寄主的木质部或韧皮部直接获取营养,而菌根异养植物则通过菌根真菌间接地获得其他植物的营养。正是由于具有这种特殊的获取营养方式,菌根异养植物一般会被错误的认为是腐生植物。本文所介绍的寄生植物仅指前一种寄生植物,即吸器寄生植物。
寄生植物的种类
根据吸器寄生植物进行光合作用的能力,可将其简单地划分为三类,即可完全光合自养的植物、只能部分光合自养的植物和不能光合自养的植物。
前两者具有叶绿体,可以进行光合作用。由于其营养不完全来源于其寄主,因此被称为半寄生植物。而不能光合自养的植物由于缺乏叶绿体,因此,其营养完全来源于寄主,故而被称为全寄生植物。
此外,后两者由于依赖于寄主的能量完成生活史,因此被称为专性寄生植物。而相应的,可完全光合自养的植物在不存在寄主的情况下也能正常生活,因而被称为兼性寄生植物。

寄生植物根据生理特性和生活史的区分
(图片来源:http://parasiticplants.siu.edu/)
寄生植物的系统发育
近年来,随着分子研究逐渐深入,目前通常认为吸器寄生植物在被子植物的演化史上独立起源了12次,分属于12目27科292属,约4750种。

12个独立起源的寄生被子植物类群的代表:A. 樟目Laurales樟科Lauraceae; B. 胡椒目Piperales鞭寄生科Hydnoraceae; C. 虎耳草目Saxifragales锁阳科Cynomoriaceae; D. 蒺藜目Zygophyllales刺球果科Krameriaceae; E. 金虎尾目Malpighiales大花草科Rafflesiaceae; F. 葫芦目Cucurbitales风生花科Apodanthaceae; G. 锦葵目Malvales岩寄生科Cytinaceae; H. 檀香目Santalales绝大部分科; I. 杜鹃花目Ericales帽蕊草科Mitrastemonaceae; J. 紫草目Boraginales沙菰科Lennoaceae; K. 茄目Solanales旋花科Convolvulaceae; L. 唇形目Lamiales列当科Orobanchaceae.
(图片来源:Nickrent, D.L. Parasitic angiosperms: How often and how many? TAXON. 2020 69(1): 5-21.)
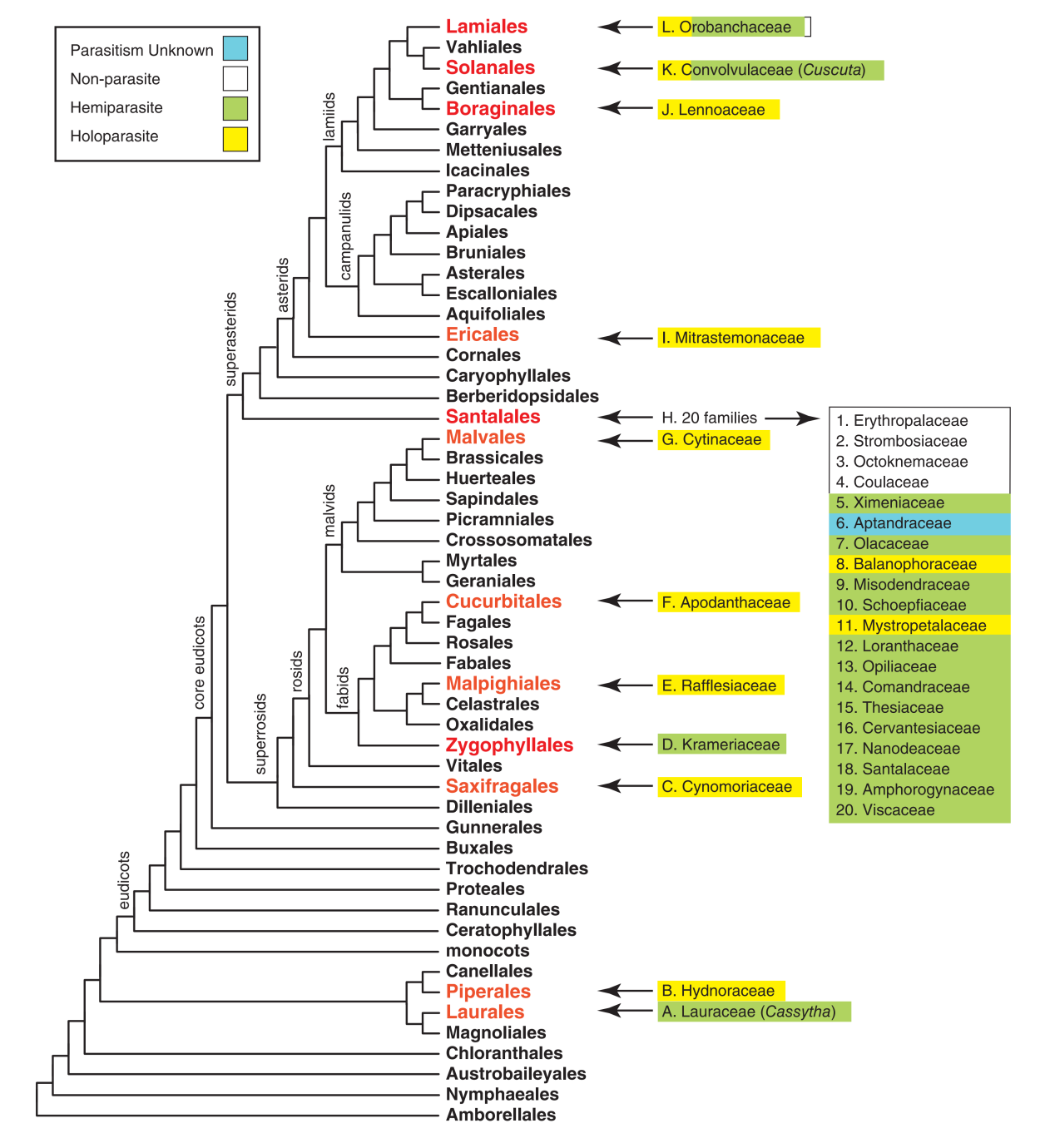
一个简化的被子植物系统发育树,显示了寄生植物所在的目红色表示含有寄生类群的目; 绿色表示半寄生类群; 黄色表示全寄生类群;白色表示自养; 蓝色表示寄生形式未明.
(图片来源:Nickrent, D.L. Parasitic angiosperms: How often and how many? TAXON. 2020 69(1): 5-21.)
在这12个类群中,半寄生的种类占多数,而全寄生的种类占少数。其中,全部为半寄生的类群只有两个——樟科Lauraceae无根藤属Cassytha和刺球果科Krameriaceae刺球果属Krameria。全部为全寄生的类群有7个,分别是鞭寄生科Hydnoraceae、锁阳科Cynomoriaceae、大花草科Rafflesiaceae、风生花科Apodanthaceae、岩寄生科Cytinaceae、帽蕊草科Mitrastemonaceae、沙菰科Lennoaceae。
既有半寄生类群又有全寄生类群的包括旋花科Convolvulaceae菟丝子属Cuscuta、檀香目Santales以及列当科Orobanchaceae。后两者恰好是最大的两个寄生植物类群。在檀香目中约有2428种寄生植物,而在列当科中,约有2163种寄生植物。
更有趣的是,这两个庞大的寄生类群的系统发育树中,在基部存在一些与寄生类群互为姊妹群的光合自养类群,这为科学家们研究寄生植物的演化打开了一扇窗口。
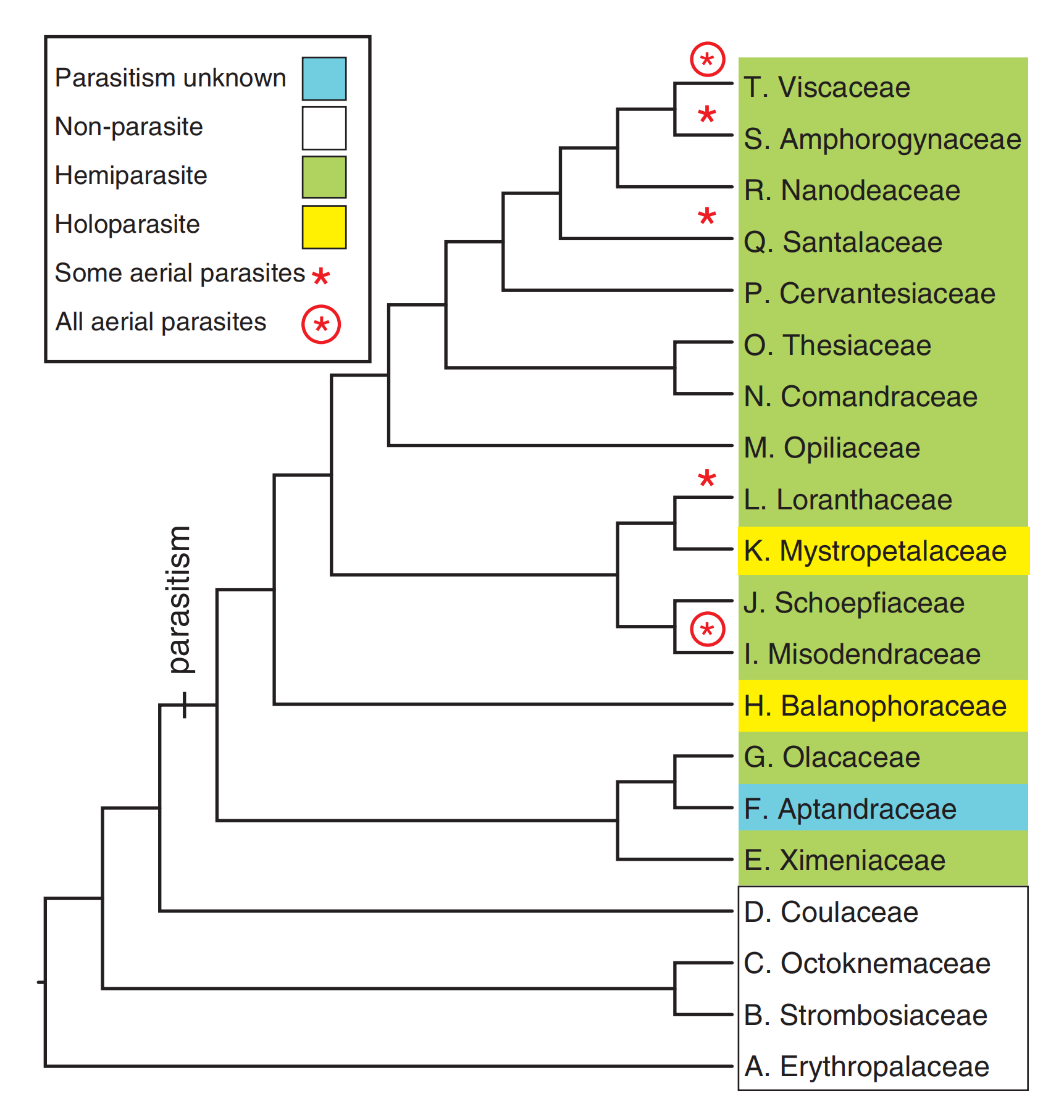
檀香目的系统发育树,共20科,可见基部4个科均为光合自养类群,剩余16个科中有13个科为半寄生,1个科寄生形式未明,2个全寄生科镶嵌在半寄生科中。
(图片来源:Nickrent, D.L. Parasitic angiosperms: How often and how many? TAXON. 2020 69(1): 5-21.)
例如,根据分子数据,檀香目共可分为20个科。最基部的4个科全部为自养类群。在剩下的16个科中,13个科为半寄生类群;仅有两个科为全寄生类群(蛇菰科Balanophoraceae,盾片蛇菰科Mystropetalaceae),并且镶嵌在半寄生类群中。
通过檀香目寄生类群以及近缘光合自养类群的系统发育关系对比,我们可以得出关于寄生植物类群起源的结论。即寄生类群是由光合自养类群演化而来的,而全寄生类群又是从半寄生类群演化而来的,且这一演化过程在檀香目中独立出现了两次。
在寄生中不断适应环境
**对于寄生植物而言,**长期依赖寄主来完成一部分生理活动,压缩了其在形态上、生理上以及基因上的进化选择。
其中一点就是性状的消失,这或许是由于在寄生生活中,维持某一性状的代价超过了其收益,或者是因为在寄生状态下,性状对适合度不再重要,因此就随着遗传漂变而丢失了。
性状减少是所有寄生植物的一个共同特征,但一旦一个关键性状丢失,寄生植物就必须依赖寄主来实现此前这个性状存在的目的。
全寄生植物通常会损失叶绿体基因组,无法独立生活。一旦寄主的种群数量下降,或者灭绝,它的寄生植物也会面临灭顶之灾。
**对于寄主来说,**当被寄生植物寄生时,可能会出现生长缓慢的情况。同时,其与菌根真菌、病原体、传粉者或食草动物之间的相互作用也会发生不利改变。此外,对干旱敏感性增加、光合作用的能力降低、后代减少、环境适应性降低等问题也会随之而来,因此寄生植物对于寄主也有选择压力。
寄生vs寄主:相互施压
一个典型的专性寄生植物从种子萌发开始,与其寄主终其一生都有着频繁的相互作用。
首先,休眠的寄生植物种子在检测到附近的寄主根所释放的化合物时,会触发种子萌发。其次,寄生体在寄主的根上会形成吸器附着。最后,寄生植物保持附着,吸收生长和繁殖所需的营养。
在这个三个过程中,双方都在相互施加选择压力。
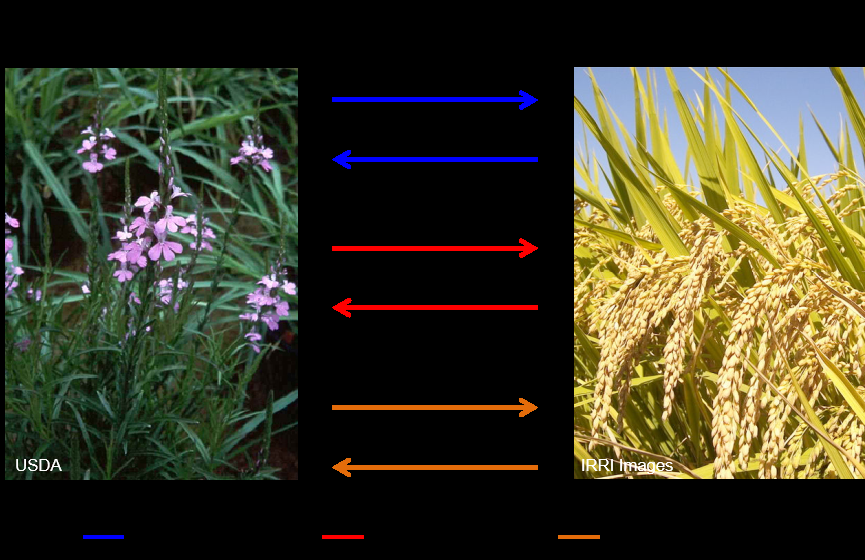
寄生植物与寄主的相互作用
(图片来源:http://parasiticplants.siu.edu/)
种子的萌发与寄主的伪装
为了保证有足够的后代,一些寄生植物可产生数万或数十万种子。这些种子通常很小,能量储备有限。在萌发时,幼苗必须在几天内附着在寄主上才能存活。
一些专性寄生植物已经进化出了特殊的发芽机制,这种机制减少了寄生植物自杀性的发芽。这些适应主要包括种子休眠和信号检测。
种子休眠通常指的是使种子在土壤中发芽延迟几十年。经过一段条件作用期,种子可以在一年中的特定时间获得发芽的潜力。
信号检测则指的是检测附近寄主根的化学信号,其中包括独角金内酯,它在极低浓度下是有效的发芽刺激剂。
而对于寄生植物与寄主而言,独角金内酯是一种必不可少的植物激素。独角金内酯是与丛枝菌根真菌(Arbuscular Mycorrhizal Fungi, AMF)建立有益的共生相互作用的重要信号。丛枝菌根真菌可以为寄主植物提供水和矿物质营养,如磷酸盐和氮,以交换来自寄主植物的碳水化合物。而独角金内酯则能够促进丛枝菌根真菌的菌丝分枝,提高其缔合效率。
能被植物所分泌的独脚金内酯有约20种,但其产生的量和种类因物种而异,甚至在物种内部也是如此。寄生植物种子可以通过对特定的独脚金内酯偏好,选择合适的寄主。
通过减少独脚金内酯的释放,寄主植物也可以对寄生植物的种子“隐形”。在农业上也能通过培育低独脚金内酯释放的作物来抵御寄生植物。
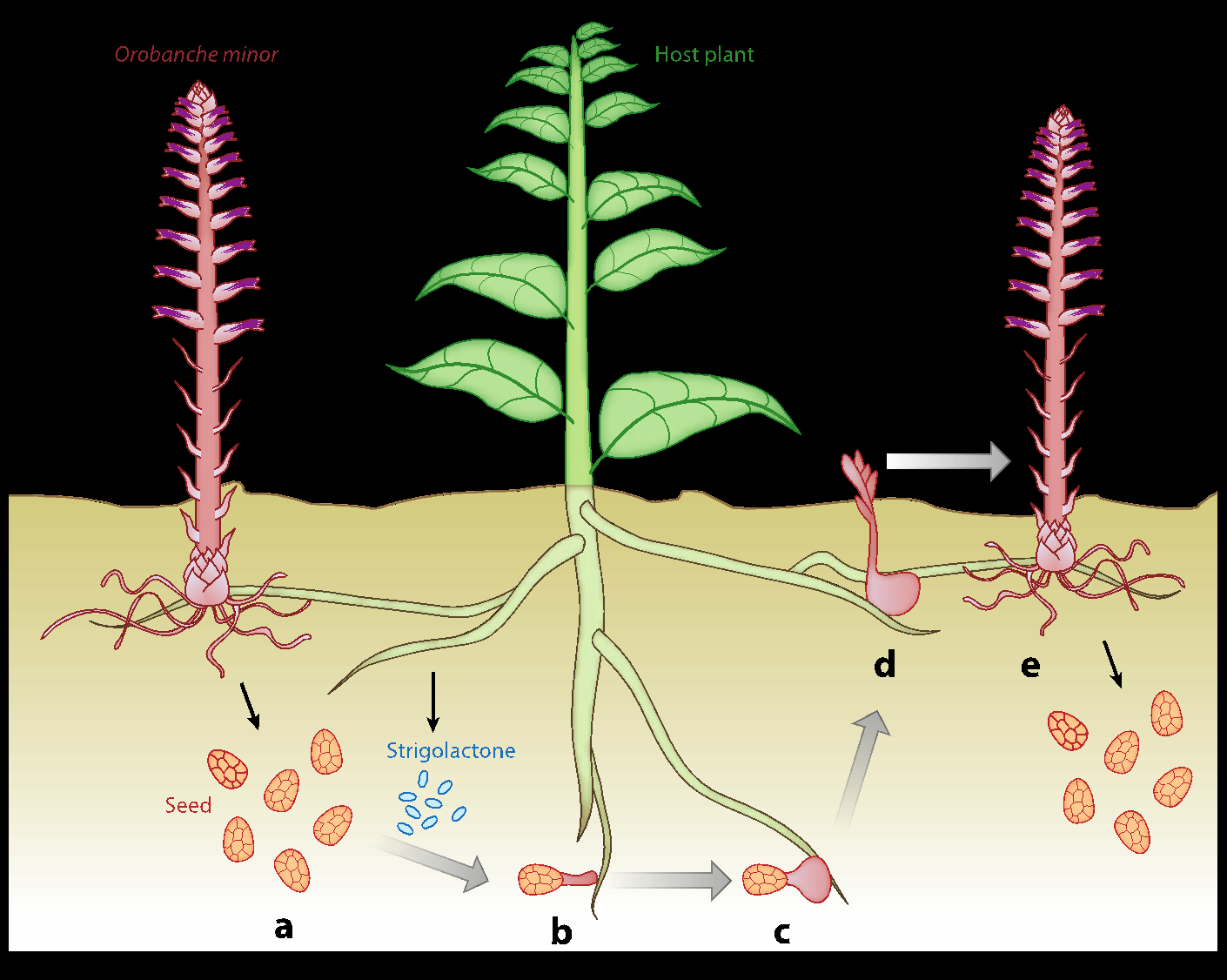
寄生植物种子通过检测寄主释放的独脚金内酯从而锁定寄主
(图片来源:http://parasiticplants.siu.edu/)
吸器的入侵和寄主的防御
当寄生植物的种子萌发,并且在寄主根分泌物中到吸器激发因子(如醌、羟基酸和类黄酮)后,寄生植物便会开始进行侵入性结构的特化。
吸器首先通过寄主根的表皮和皮层侵入寄主,然后在寄主和寄主之间形成木质部连接,寄主一般通过物理手段以及化学方式对吸器进行拦截。
通常在某些植物的根部中,坚硬的内皮层可以防止吸器的进一步入侵,而根部的酚类、植物抗毒素和其他分泌化合物可延缓吸器的发育或侵入,最终达到防御目的。
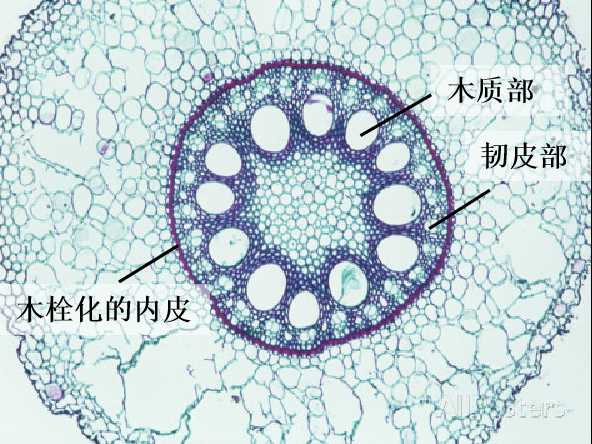
玉米木栓化的内皮可以保护木质部不受寄生植物的侵袭
(图片来源:http://parasiticplants.siu.edu/)
寄主的“反抗”——毒害作用
在连接到寄主的木质部(有时是韧皮部)后,植物就开始吸取寄主的营养。专性全寄生植物必须保持与寄主的连接来生长繁殖,完成它们的生命周期。
维持与寄主根系持续且畅通的维管组织连接的能力,是刺激寄主反应的重要前提。有的寄主已经能够做出防御反应,它们通过阻塞寄主本身的维管组织来关闭与寄主植物的维管连接,或是产生可以在整个寄生植物体内扩散的有毒化合物。
此外,通过干扰寄生植物体内的维管组织分化也是部分寄主所做的防御反应。目前已经在一些玉米和向日葵野生品种中发现了相关的抗性基因,这些抗性基因能够有效阻止寄生植物发育,在农业上有着重要用途。
被误解的寄生植物
在农业和林业上,一些寄生植物由于其影响寄主正常生长发育的特点,成为一类“有害植物”。但实际上,这些植物虽然会损害农林作物,他们也是当地生态系统的重要基石。寄生植物对生物之间的竞争关系、物质循环、能量流动、信息传递以及生态系统的多样性有着重要的积极作用。
除此之外,更多寄生植物相关的分子形态学研究,尤其是对于寄生植物及其近缘自养类群的演化研究正在逐步为我们揭示系统发育学以及进化生物学上的关键问题,为现代分类学的发展打下基础。
结语
或许有些人会因为寄生植物的特性而欲加之罪,但站在寄生植物们的角度而言,它们不过是在选择的压力下走出了一条另辟蹊径的道路。目前,许多寄生植物正遭遇着濒临灭绝的风险,如蛇菰、肉苁蓉、锁阳等,因其植株形状独特而被追捧为所谓的“回春草”、“壮阳草”,正被大肆挖掘,这些植物的种群受到严重威胁,亟待我们的重视保护。
编辑:应奕可
参考文献:
【1】Martin et al. Symbiosis: 'Living together' in chaos, Studies in the History of Biology. 2012. 4 (4): 7-25.
【2】Poulin, R. Rollinson, D. The Many Roads to Parasitism: A Tale of Convergence. Advances in Parasitology. Academic Press. 2011. 74: 27-28.
【3】Dörr, I. How Striga Parasitizes its Host: a TEM and SEM Study. Annals of Botany. Oxford University Press. 1997. 79(5): 463-472.
【4】Bromham, L., Cowman, P. & Lanfear, R. Parasitic plants have increased rates of molecular evolution across all three genomes. B. M. C. Evol. Biol. 2013 13: 126.
【5】Su et al. Novel genetic code and recordsetting AT-richness in the highly reduced plastid genome of the holoparasitic plant Balanophora. Proc. Natl. Acad. Sci. U.S.A. 2019. 116: 934-943.
【6】Yang et al. Horizontal gene transfer is more frequent with increased heterotrophy and contributes to parasite adaptation. Proc. Natl. Acad. Sci. U.S.A. 2016. 113: E7010-E7019.
【7】Hatcher, M.J., Dick, J.T. & Dunn, A.M. Diverse effects of parasites in ecosystems: Linking interdependent processes. Frontiers Ecol. Environm. 2012 10: 186-194.
【8】Wang, Y. & Bouwmeester, H.J. Structural diversity in the strigolactones. J. Exp. Bot. 2018. 69: 2219-2230.
【9】Nickrent, D.L. Parasitic plants. in: McGraw-Hill Yearbook of science and technology. New York: MacGraw-Hill. 2008. Pp. 251-253.
【10】Nickrent, D.L. Parasitic angiosperms: How often and how many? TAXON. 2020 69(1): 5-21.
【11】APG IV. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016. 181: 1-20.
【12】Nickrent, D.L. Malécot, V. Vidal-Russell, R. Der, J.P. A revised classification of Santalales. TAXON. 2010. 59 (2): 538-558.
【13】Nickrent, D.L., Anderson, F. & Kuijt, J. Inflorescence evolution in Santalales: Integrating morphological characters and molecular phylogenetics. Amer. J. Bot. 2019. 106: 402-414.
【14】Su, H.-J., Hu, J.-M., Anderson, F.E. & Nickrent, D.L. Phylogenetic relationships of Santalales with insights into the origins of holoparasitic Balanophoraceae. TAXON. 2015. 64: 491-506.
【15】Yu, W., Randle, C.P., Lu, L., Wang, H., Yang, J., dePamphilis, C.W., Corlett, R.T. & Li, D. The hemiparasitic plant Phtheirospermum (Orobanchaceae) is polyphyletic and contains crypticspecies in the Hengduan Mountains of Southwest China. Frontiers Pl. Sci. 2018. 9: 142.
【16】Watling, J.R., Press, M.C. Impacts of Infection by Parasitic Angiosperms on Host Photosynthesis. Plant biology. 2001. 3: 244-250.
【17】Xie, X., Yoneyama, K., Yoneyama, K. The Strigolactone Story. Annual Reviews of Phytopathology. 2010. 48: 93-117.
【18】Fernández-Aparicio, M., Flores, F., Rubiales, D. Recognition of root exudates by seeds of broomrape (Orobanche and Phelipanche) species. Annals of Botany. 2009. 103: 423-431.
【19】López-Ráez, J.A., Pozo, M.J., García-Garrido, J.M. Strigolactones: a cry for help in the rhizosphere. Botany. 2001. 89: 513-522.
【20】Xie, X., Yoneyama, K., Kisugi, T., Uchida, K., Ito, S., Akiyama, K., Hayashi, H., Yokota, T., Nomura, T., Yoneyama, K.Confirming Stereochemical Structures of Strigolactones Produced by Rice and Tobacco. Molecular Plant 2013. 6(1): 153-163.
【21】Jamil, M., Rodenburg, J., Charnikhova, T., Bouwmeester, H.J. Pre-attachment Striga hermonthica resistance of New Rice for Africa (NERICA) cultivars based on low strigolactone production. New Phytologist 2011. 192: 964-975.
【22】Westwood, J.H., Yoder, J.I., Timko, M.P., dePamphilis, C.W. The evolution of parasitism in plants. Trends in Plant Science 2010. 15: 227-235.
【23】Amusan, I.O., Rich, P.J., Menkir, A., Housley, T., Ejeta, G. Resistance to Striga hermonthica in a maize inbred line derived from Zea diploperennis. New Phytologist 2012. 178: 157-166.

来源: 中国科普博览
内容资源由项目单位提供
