
 科普中国公众号
科普中国公众号
 科普中国微博
科普中国微博

 帮助
帮助
 中国微生物学会
中国微生物学会 1. 引言
病毒,这些微小的生物实体,形态与类群多样,与人类的生活息息相关。它们几乎分布于地球上的各种环境,研究人员在深海、热泉、盐湖和极地生境均发现了病毒。病毒既是许多疾病的元凶,也是生物进化的重要推手。人们难免会产生疑问,病毒从哪里来?病毒是如何演化的?为什么病毒这么容易变异?本文将带你走进病毒的起源世界,探索这些神秘生物的起源之谜。
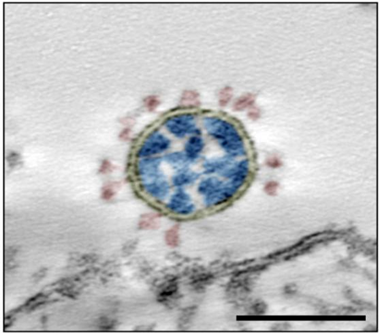
图1. 新冠病毒电镜图
注:黄色:病毒包膜;红色:刺突蛋白(S蛋白);蓝色:核衣壳蛋白及核酸。比例尺 = 100 nm。(图片来源:J, Nitsche A. Sci Rep. 2021; 11(1): 3515)
2. 病毒的起源假说
根据生物演化的理论,科学家将所有生物所产生的演化总结成一棵生物演化树。有理论认为现代细胞起源于同一个祖先——LUCA,但由于病毒极其微小且没有硬质结构,不易形成化石,导致基本没有化石证据能够证明病毒的出现与其演化过程。目前只能通过现有的病毒做分子生物学测定来推测病毒的演化过程。病毒结构较为简单,为蛋白质外壳包裹着RNA或DNA构成的核衣壳结构,比较复杂的病毒还含有包膜结构与脂类、多糖、蛋白质或多胺类等物质。病毒无法独立完成复制并形成子代病毒,必须通过侵染细胞来完成自身的生命周期。目前常见的假说包括以下几种:
2.1 自然发生说
有学者提出复杂的化学反应带来了有机大分子,这些大分子中的DNA、RNA及蛋白质等开始相互独立或者彼此协作地复制自身,形成“亚生命单位”,再进化成为亚病毒或者病毒。一些病毒或许是在环境的压力下想要保全自身,或许是想要以压倒性的优势战胜其他的竞争者,与脂质多糖等分子相结合形成细胞的雏形,而病毒成为了细胞与大分子之间的“过渡类群”。有学者根据细胞演化过程推测真核细胞病毒可能是由古菌病毒演化而来,这种理论也被称为病毒自起源理论。这种理论曾经一度十分盛行,毕竟病毒的结构是如此简单,它们与细胞生命的差异也非常显著。然而,这种理论却随着分子生物学研究的不断深入而被质疑。
2.2 共进化假说
病毒起源的第二种假说主张病毒起源于细胞。现代科学认为病毒可能与宿主细胞共进化。病毒可能起源于宿主细胞中的某些遗传元件,如转座子,这些元件在特定条件下能够获得自我复制的能力,最终演化成病毒。某些原核生物中的质粒为可在细胞间传递的遗传物质。病毒与质粒的生物学属性具有相似性。比如大肠埃希氏菌病毒P1,进入大肠杆菌后如果启动复制周期,表现出病毒的特征;反之,显现质粒的特征。此外,并非只有原核生物才会出现该种情况,在人体细胞中也分布一类核酸序列——转座子。根据分子生物学检测,很多转座子都有着和病毒非常相似的基因序列,它们将自己整合到宿主细胞染色体上的机制也和病毒出奇相似,尤其是其中的“逆转录转座子”与一些逆转录病毒相似度极高。今天,我们依然可以在人类染色体上找到曾经整合到人类染色体中的病毒片段——“内源性病毒”。
2.3 退化假说
另一种观点是“退化假说”,认为病毒可能是从较大的生物体中退化而来的。这一理论的支持者认为,病毒可能是早期生物体的一部分,随着时间的推移,它们逐渐失去了独立生存的能力,转而依赖宿主细胞生存。2003年科学家发现了一种巨型病毒,被称为“拟菌病毒”,体型达到了0.4 - 0.5μm。拟菌病毒的基因序列与古菌高度相似,不同的是拟菌病毒丢失了一部分完成自我复制的关键基因。2008年科学家又发现了另一种体型“巨大”的病毒,称为“米米病毒”。
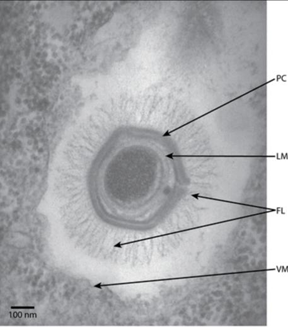
图2. 米米电镜图(图片来自:Claverie JM, etal. Annu Rev Genet. 2009; 43: 49-66)
3. 结语
病毒的起源是一个复杂且引人入胜的话题。从自然发生说到共进化假说,再到退化假说,科学家们一直在探索病毒的起源之谜。也许,病毒的起源远比这些假说更为复杂,甚至未必是唯一的来源,但是病毒却真真切切地在我们的“演化中”留下了深深的烙印。病毒与宿主之间的相互作用、病毒的高突变率、基因重组以及跨种传播,都是病毒进化的重要方面。了解这些知识,不仅有助于我们更好地理解病毒,也为我们提供了防控病毒性疾病的科学依据。
作者简介:朱宏莉,博士、教授、博士生导师,西北大学生命科学学院生物技术系主任,中国生物医学工程学会血液代用品分会副秘书长,中国微生物学会会员,陕西省微生物学会常务理事。
主要参考文献:
1.Abrahao J, Silva L, Silva LS, et al. 2018. Tailed giant tupanvirus possesses the most completetranslational apparatus ofthe known virosphere. Nature Communications, 9(1): 749.
2.Gudbergsdottir SR, Menzel P, Krogh A, et al. 2016. Novel viral genomes identified from sixmetagenomes reveal wide distribution of archaeal viruses and high viral diversity in terrestrialhot springs. Environmental Microbiology, 18(3):863-874.
3.冯元昱, 董昊东. 病毒自起源理论的研究进展. 生物化工. 2019, 5(03): 137-139.
4.GuyL, SawJH, Etema TJ. The Archaeal Legacy of Eukaryotes a Phylogenomic Perspetive. ColdSpring HarbPerspect Biol, 2014, 6 (10) : a016022.
5.李赞, 李绍军. 病毒与细胞在起源与进化上的关系. 生物学教学. 2012, 37(10): 4-6.
6.Chuong EB, Elde NC, Feschotte C. Regulatory activities of transposable elements: from conflicts to benefits. Nat Rev Genet. 2017; 18(2): 71-86.
7.安亚龙, 陈子璇, 迟诚林, 陈才, 宋成义, 王宵燕. 动物反转录转座子研究进展. 农业生物技术学报. 2021, 29(01): 125-136.
8.Xiao C, Chipman PR, Battisti AJ, et al. Cryo-electron microscopy of the giant Mimivirus. J Mol Biol. 2005; 353(3): 493-496.
9.Schulz F, Roux S, Paez-Espino D, et al. Giant virus diversity and host interactions through global metagenomics. Nature. 2020; 578(7795) : 432-436.
10.Claverie JM, Abergel C. Mimivirus and its virophage. Annu Rev Genet. 2009; 43: 49-66.
来源: 陕西省微生物学会
