最近,就读于某植物研究所的准硕士三年级研究生小美有点苦恼。她茶饭不思,甚至连实验室的离心机都没心情抢。原来,前一天晚上,隔壁实验室的小帅向她表白了,还约她三天后去吃烧烤(即约会)。
小美对小帅不太熟,实验室毕竟隔墙如隔山。她只能通过自己严谨的思维与全方位的综合考虑,做出“接受”OR“拒绝”的决定:
接受→她可能不得不放弃宝贵的实验时间。众所周知,小美马上就研三了,是人生又又又又一个关键时期,稍有延误,实验无法按时完成、文章没法接收、延期毕业、不能按时考博、博士不能按时毕业,最终找不到工作、失去年龄优势、申请不到基金、退休前评不到正高,完啦!科研道路全毁了!
拒绝→她可能没法在30岁前找到对象、35岁前生不了孩子、孩子没法在她退休前独立,完啦!人生规划全毁了!况且,不能落入经验主义的圈套中,要用理性和发展的眼光看问题,维持现状并不一定是最好的。万一小帅实验技能卓越,拒绝他岂非错失一台多功能全自动“点样机”?
小美踌躇不已,然而三天内就要做出回复。于是她决定采取一种古老但科学有效、同时符合她植物学研究生身份的方法来做决定:数花瓣——单数接受、双数拒绝。
小美快速地制定了实施方案,唯一的问题是:要用什么植物的花来数呢?
首先,要明白“数花瓣”这件事的本质——“花瓣数量的奇偶性”。
植物的花瓣数量是否有规律呢?在回答这个问题之前,先明确一下“花瓣”是什么?
裸子植物没有真正的花,花是被子植物的生殖器官。一朵比较典型的花可以分成几个部分,除了花梗和花托,从外到内直观的有:花萼、花冠(单个花瓣称为花瓣,所有花瓣的总和称为花冠)、雄蕊群和雌蕊群。这些花部分之间有时候泾渭分明,有时候又模糊不清。

有时花萼和花瓣难以区分,索性也就不区分,直接合称为花被片;有时候花瓣会长成一桶像连衣裙的东西,基部连在一起,称为合瓣花,顶上分开的部分,称为裂片;有时候我们以为的花瓣并不是花瓣,而是其他的东西,比如说三角梅的“花瓣”其实是苞片;有时候花瓣数量很难说清,比如说向日葵是一堆小花长在一起拼成的一个头状花序,如果统计小花的花瓣数量,那么向日葵周围每朵小花只有一片花瓣(舌状花)、内部的每朵小花有4-5个裂片(管状花),我们是数哪种小花的花瓣数量?抑或数整个花序的花瓣数量?




为了避免小帅过后申请复查,数花瓣就要数地地道道的花瓣,哪怕小帅找公证处来也只能承认是花瓣:1)合瓣花,数裂片数量;2)无法明确区分花瓣花萼、但是看上去像花瓣的(柔软、非绿色、光滑、不像叶子),全当花瓣数;3)如果花瓣和花萼有区分,但是中间有过渡状态,这种花不用于计数;4)三角梅和向日葵这样“暧昧”的花不用于计数;5)像水稻那样,花瓣退化或缺失而难以辨认的花不用于计数(没人想数水稻花吧)。
技术路线A:自然界有规律的花瓣数量
花是进化来的。起初,植物并没有花,也没有花瓣,后来叶子发生了变异,形成了花瓣,花瓣数量也是植物进化的结果。因此花萼、花瓣、雌雄蕊的着生方式同叶的着生方式有异曲同工之处——被子植物叶片有螺旋和轮生等着生方式,花器官也有螺旋和轮生。要注意的是,螺旋花不一定所有部分都是螺旋的,比如说木兰科,它们的雄蕊螺旋着生,但花被片轮生;睡莲目的花瓣螺旋着生,但萼片常常只有1轮(3-5枚左右)。目前还说不清到底是螺旋花更原始还是轮生花更原始。

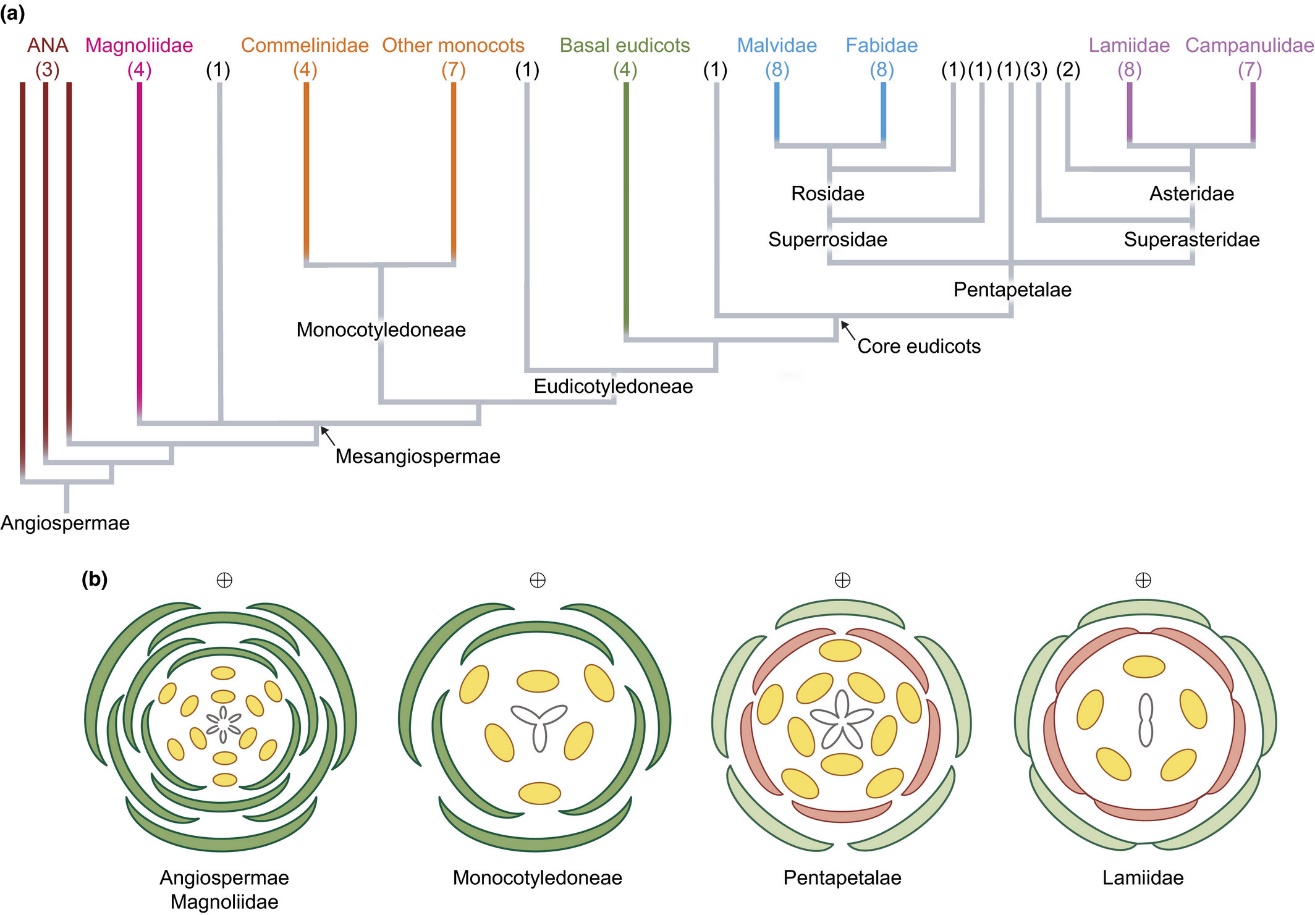
(a)简化的被子植物生命之树,分支顶部括号中的数字是该分支包含的“目”(order)的数量,注意将分支的名称同下面(b)中的名称对应起来;(b)中的示意图被称为“花图示”,能很清楚地反映出花内各器官的排列方式:最中间的是雌蕊(心皮),黄色的圆点是雄蕊,雄蕊外面是花被片,如果花被片无法区分,则花萼和花瓣都画成绿色,如果可以区分,那么内圈红色的是花瓣,外圈绿色的是萼片(Smyth, 2018)
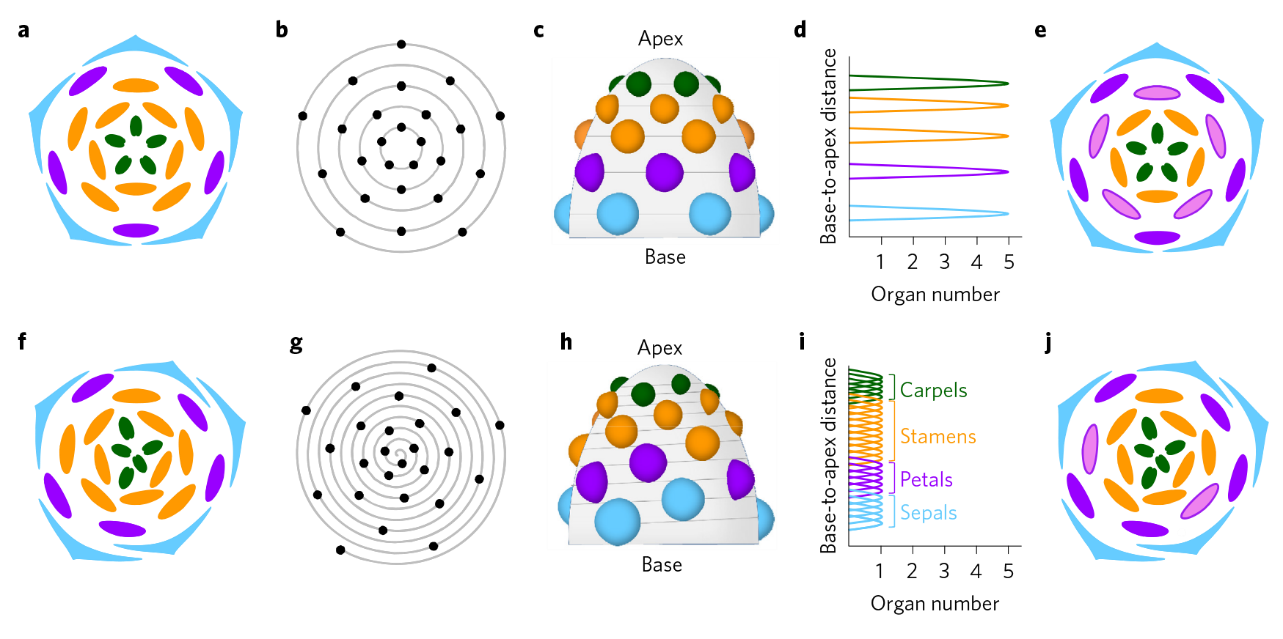
上排:轮状花,下排:螺旋花;a和f:花图示;c和h:花器官在花托上的空间分布示意图;d和i:花器官数量沿花基部至顶端轴的变化;e和j:第三轮雄蕊瓣化时的花图示(Wanget al., 2015)
(一)螺旋花
螺旋花的花瓣数量往往符合一个神奇的数学规律——斐波那契数列,又称黄金分割数列,相邻两个花器官(如花瓣)之间形成137.5°的斐波那契角。据说是因为这样排列叶片和花瓣不容易相互重叠,植物可以用更高的性价比获得充足的阳光和花朵展示机会。
所谓斐波那契数列,就是从0、1开始,每个数字是前两个数字的和,具体就是:0、1、1、2、3、5、8、13、21、34……由于这个数列开局是0和1,也就是一个偶数和一个奇数,偶数+奇数必然是奇数,所以第3个数一定是奇数;奇数(第2个数)+奇数(第3个数)=偶数(第4个数);奇数(第3个数)+偶数(第4个数)=奇数(第5个数)……总结一下,斐波那契数列的奇偶性排列一定是:偶奇奇偶奇奇偶奇奇……也就是“偶奇奇”循环。这还了得,奇数比偶数多!

螺旋着生的叶和花器官往往符合斐波那契数列。(A)云杉的螺旋叶序;(B)番茄顶芽;(C)非洲菊头状花序;(D)原基分化早期(Reinhardt & Gola, 2022)
然而这不意味着自然界中花瓣数量奇数的概率一定会更大,举个例子:假设全世界只有四种花瓣数量类型——1、2、3、5,显然,奇数比偶数多,但是花瓣数量=2的植物有几万种,其他花瓣数量的植物只有几千种,那么哪怕“奇数阵营”多样性高,它们在数量上成不了主流。因此需要考虑**“花瓣数量”这个性状的数据分布**。这是个庞大的问题,小美不一定能在三天内查到所有数据,然而她可以采取化整为零的策略——螺旋花植物也没多少,把有代表性的类群拉出来数数即可。

花器官发育顺序。(a-c)轮生花,轮生花的花原基早起发育的时候也可能是螺旋开始的,可以连出斜列线(parastichies)和直列线(orthostichies);(d-f)螺旋花,只有斜列线没有直列线,螺旋花和轮生花的发散角不同(Endress & Doyle, 2007)
花瓣螺旋着生的类群包括:五味子科(Schisandraceae)、苞被木科(Trimemiaceae)、蜡梅科(Calycanthaceae)、香皮茶科(Atherospermataceae)、毛茛科(Ranunculaceae)部分类群、无油樟属(Amborella)、八角属(Illicium)、木兰藤属(Austrobaileya)、莲属(Nelumbo)、单心木兰属(Degeneria)、奇子树属(Idiospermum)、奎乐果属(Gomortega)、八角桂属(Hortonia)……
这都是些什么佶屈聱牙的名字?!其实这些类群中很多不分布在我国,如毛茛科中典型的螺旋花黑种草属(Nigella)主要分布在欧洲,我国只有两个引入观赏的栽培种,而且黑种草属看上去很像花瓣的部分其实是萼片,真正的花瓣藏在萼片和雄蕊之间,萼片和花瓣均以5瓣为多。
在这一堆螺旋花植物中,大部分人比较熟悉的有蜡梅和八角,如果解剖一下,会发现它们的花瓣数量确实不稳定(符合我们的要求),但是数起来有点费劲——苞片、萼片、花瓣之间有过渡状态,不容易区分,不符合小美的计数规则。五味子科植物的花被片花瓣状,5-24枚,数量也不确定,这似乎挺好,但是五味子还要上山去找,不太方便。
莲花(Nelumbonucifera)是个不错的选择,不同品种的花瓣数量差异非常大,比如说“金秋”124瓣左右,“白花建莲”16瓣左右,其杂交后代花瓣数量奇偶性无显著差异。野生荷花花瓣数量在15-22之间,只要选对了莲花品种或野生居群,不难roll出花瓣数量奇偶性均衡的数花瓣材料。
考虑到时间紧、预算少,先暂时把莲花列入备选名单中。至于苞被木、无油樟之类就不考虑了,总不能为了数个花瓣数量申请标本传递吧?小帅何德何能?

白色分支主要是螺旋花被片、黑色分支主要是轮生花被片,其余介于两者之间。轮生花被片不仅占据的分支多,分支内包含的物种总数更多(Endress & Doyle, 2007)
(二)轮生花
自然界中轮生花占大多数,其花瓣数量往往符合某种“花基数”,即一轮有n个花瓣,称为n基数。常见的花基数包括:2、3、4、5、6,甚至7基数以及更高。一朵花可以有很多轮花瓣,最终花瓣数量就是n的倍数。显然,轮状花的花瓣数量由基数和轮数共同决定。常见的花基数和轮数是多少呢?

单子叶植物花主要为3基数,经常有两轮,即6枚花瓣。双子叶植物花主要为4、5基数,以5基数为多,很少有两轮成10枚花瓣(野生的杜鹃花偶尔会突变,长出两轮花瓣,形成10个花瓣裂片)。4基数可能是5基数花丢失一个基数导致的,2基数则有可能是3基数花丢失了一个基数导致的。6基数及以上的花比较少见。
常见植物,比如说葱、百合、鸢尾、鸭跖草等等是3基数花,双子叶植物中的木兰科也是3基数花;桃、李、梅、樱、杜鹃、海棠、蔷薇、蜀葵、石竹等等是5基数花;桂花、油菜花、虞美人等等是4基数花。为什么3和5这两个数字这么特别?有人认为可能是由于3和5刚好是斐波那契数列中的数字,如果轮状花是从螺旋花进化来的,花基数倾向于斐波那契数列中的数字似乎说得过去,但是这个观点尚未获得证实。
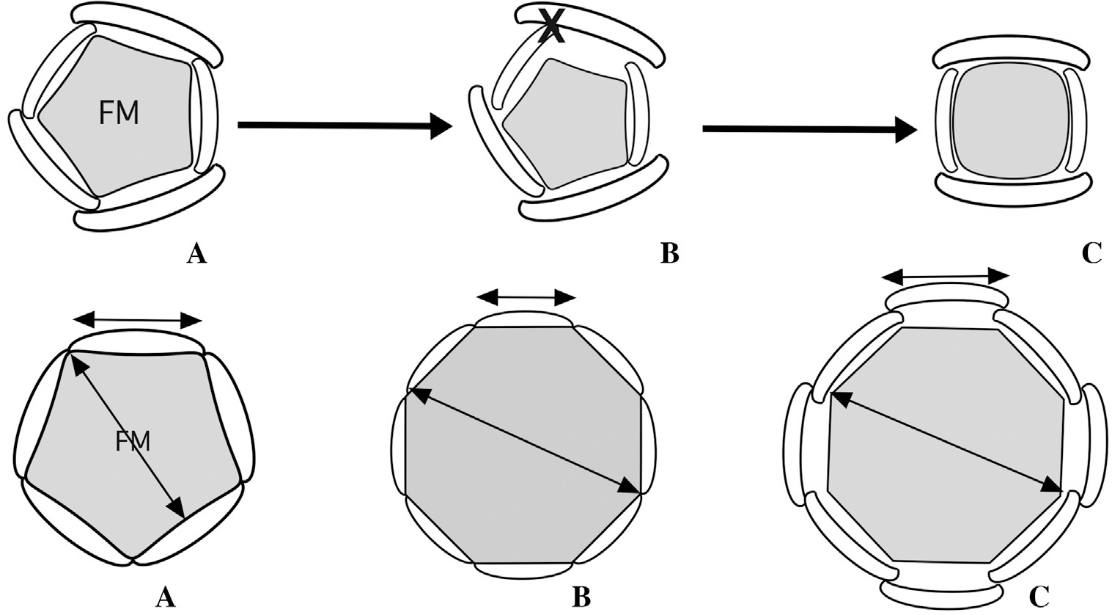
上排:(A)5基数花,(B)花分生组织缩小导致发育空间减少,丢失一个花器官,(C)4基数花;下排:(A)稳定的5基数花,(B)花分生组织增大、花器官减小,导致花基数增加而轮数不变,(C)花分生组织增大、花器官也增大,放不下这么多花器官,所以大家只能“错身站着”,导致花器官重新排列,轮数增加,A和B都可能会产生C(De Craene, 2016)
总结来说,如果小美随便在被子植物名录里选一种植物来数花,那么她大概率得出的结果是奇数。瞧瞧!这还有公平性可言吗?不如别数,直接接受小帅得了。别怕,我们还有Plan B。








技术路线B:栽培品种的不规则花瓣数量
同一种植物的花瓣数量并不是不变的,4瓣的植物偶尔也会长出5个花瓣,生长环境以及随机等因素都有可能导致植物的花瓣数量发生波动,比如说低温可以让栽培的重瓣月季花长出更多花瓣、植物激素(如赤霉素和生长素)让凤仙花和香石竹花瓣增多等等。另外,花瓣越多、变数越多,对于螺旋花来说,如果你长了10圈花瓣,似乎再多长一两圈也很正常;对于轮状花来说,花基数越大,越不稳定。
人类在培育观赏花卉品种时,会刻意选育一些花瓣多的品种,看上去雍容富贵。人类势力的加入,给花瓣数量带来了更多不确定性。为啥人工选育能弄出这么多花瓣?导致花瓣数量增加的原因很多,包括积累起源(主要针对离瓣花)、苞片起源(苞片或花萼变成类似花瓣的器官)、雌雄蕊起源(雌雄蕊瓣化)、台阁起源(正常开放的花上层又开出一朵花),重复起源(花内层结构重复外层的结构和基数)和花序起源(针对单花组成的花序,如让菊花外轮多圈小花变成舌状小花)等,其中最重要的一种是:雌雄蕊瓣化。
雌雄蕊瓣化的底层原理比较复杂,这要从花瓣分化的本质,也就是基因调控说起——ABC模型,以及它的升级版ABCDE模型,其中研究最多的重瓣相关基因是C类和A类基因。简单来说,就是某些基因突变后,在该长雄蕊的地方长出了花瓣、该长雌蕊的地方长出了花瓣,花瓣自然就变多了。染色体加倍等其他原因也可以产生重瓣花。另外,还有些基因突变可以改变花基数。
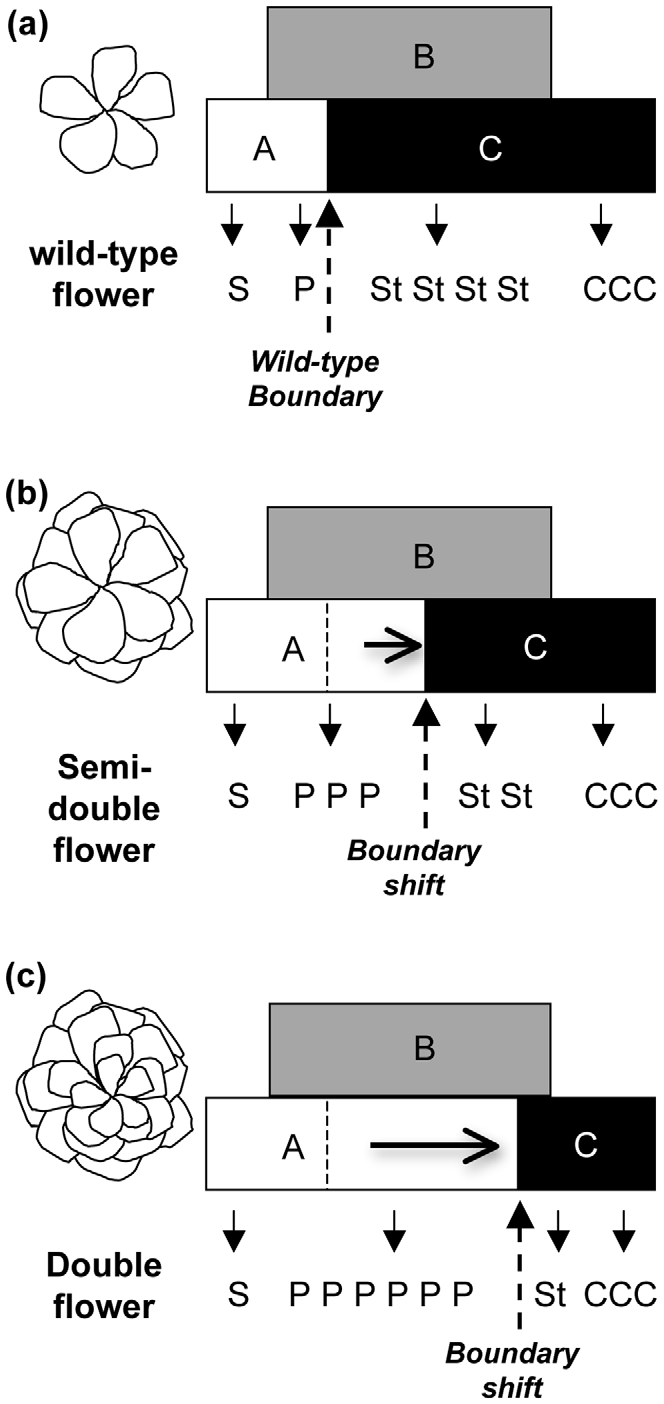
ABC模型对月季重瓣花形成的解释。(a)野生型只有一轮花瓣;(b)半重瓣花和(c)重瓣花的花瓣和雄蕊边界不稳定(Dubois et al. 2010)
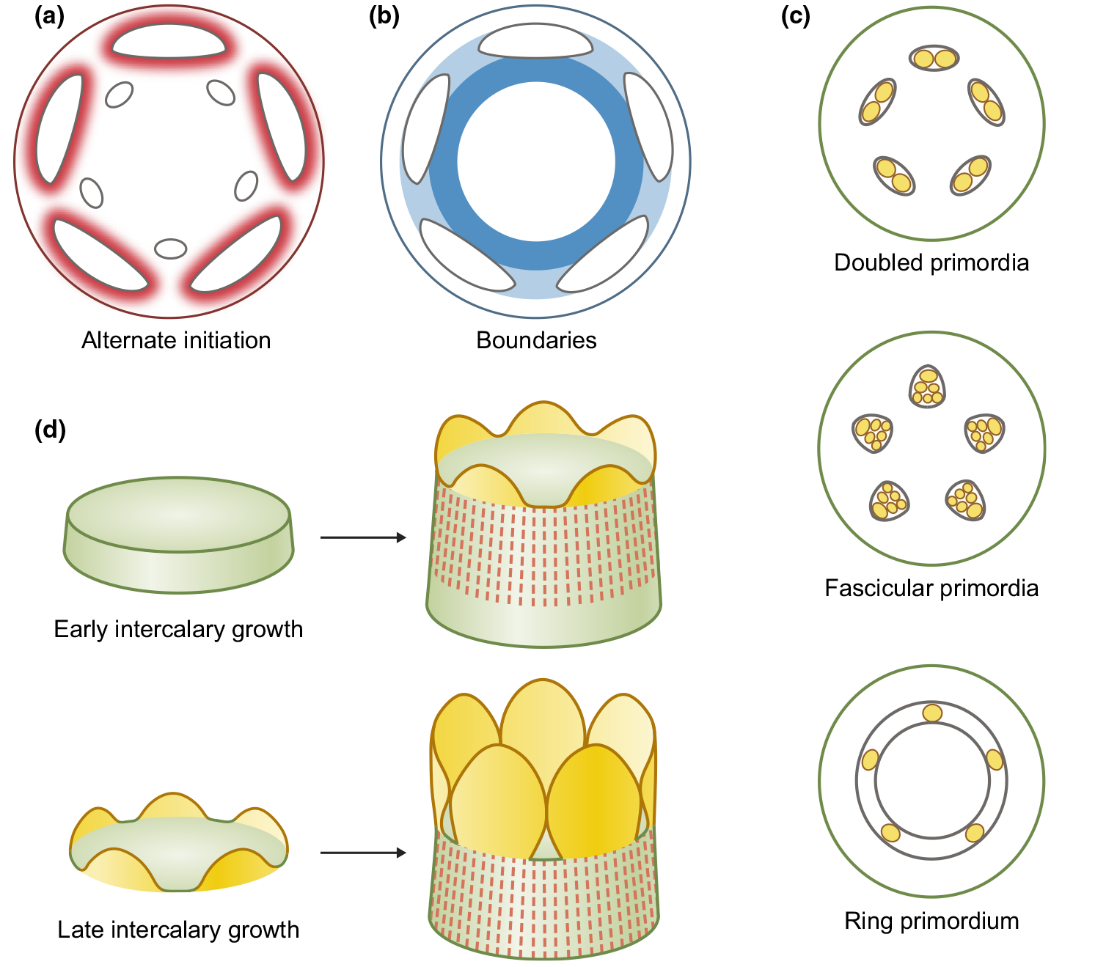
花器官排布的机制。(a)红色表示生长素浓度,越高则越红、对花器官分化的抑制作用越强,所以新的原基间隔着产生(白色的小椭圆形);(b)深蓝色表示(a)图中新原基的边界;(c)雄蕊分化的时候经常在一个初始原基内部产生多个小原基(黄色的圆圈);(d)花器官基部融合的不同情况:花器官发育早期融合(上)和花器官发育一段时间后从原基下方发生融合(下)(Smyth, 2018)



小美又有话要说了,重瓣不就是加倍了吗?无论是几基数的花,两轮就变成偶数了。一个品种,基数是确定的、重瓣的轮数是确定的,只要她选定了品种,那相当于直接选择了花瓣数量的奇偶性,这还数什么?其实,重瓣有很多类型,不一定是整倍的,瓣化程度从低到高为:(1)单瓣→(2)半重瓣(花瓣层数少,保留较多正常雄蕊或雌蕊,仍然可育)→(3)牡丹型重瓣(结构松散,中心保留部分未完全瓣化的雄蕊)→(4)玫瑰型重瓣(雄蕊完全瓣化,不可育)。很少有研究会把重瓣花具体的花被片数量统计表公布出来,但是同一栽培环境下同一品种的花被片数量似乎是有规律的,比如说东方百合的3个品种最大出现频率的正常花被片数量分别为:“Siberia”最多为5或6片、“Sorbonne”最多为5片、“Fastrada”最多为6片。
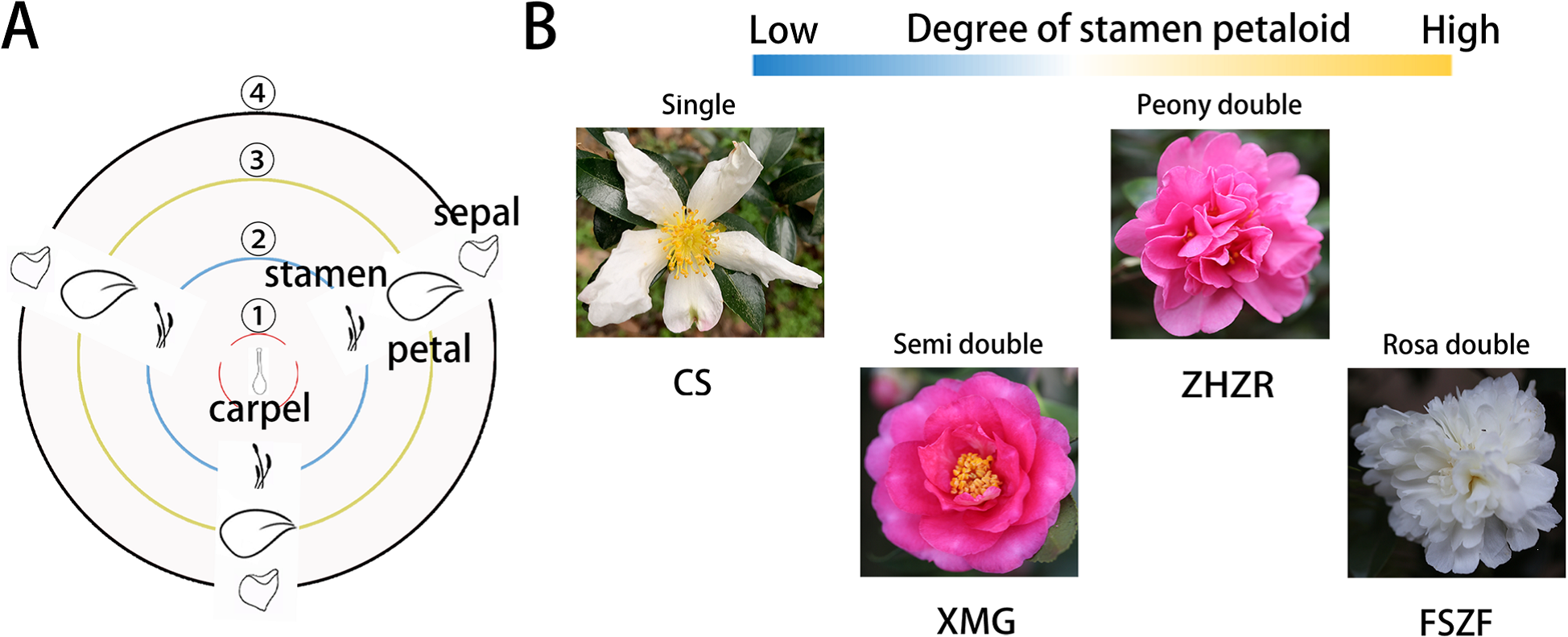
山茶花单瓣至重瓣的瓣化程度(Fan et al., 2022)



啊!天哪!数个花瓣怎么这么复杂!我们已经就这个问题讨论4千多字,快赶上一篇小论文的字数了!不如随便选一种花瓣奇偶不确定的花,先买来统计下,如果奇偶数分布是对称的(即奇数和偶数概率相同),那我们就正式用这种花的花瓣数量来决定是否去约会,如果奇偶数不对称就换一个品种,直到满足我们的要求。
栽培的重瓣月季花/玫瑰花就是很好的选择!同一个品种“月月粉”有的研究中数出来是23片花瓣,有的研究数出来是20片花瓣。当然,如果小美有闲情也可以去大观公园趴栏杆上数几天荷花,显然她没什么闲情。
现在我们可以敲定最终方案:1)网购35朵月季花(玫瑰花);2)用其中30朵统计出花瓣数量的奇偶性分布,如果花瓣数量奇、偶数出现的概率显著偏离0.5,就重新购买一批月季花,直到该批次奇偶性呈现出比较均匀的分布;3)使用剩下的5朵花统计花朵数量奇偶性,如超过3朵为奇数,就选择接受小帅的邀请,反之就拒绝。
小美舒了口气:不愧是我,简直滴水不漏,希望小帅可以耐心等待一批奇偶均匀的月季花到货。
果然,知识改变命运。
【完】
感谢读到最后,这是“室花小美的脱单之路”系列的第一篇(不保证有第二篇)。植物的花瓣数量是一个有趣的研究方向,有各种不同的研究角度,比如说:调控基因、生理代谢过程、花原基发育、花瓣数量在整个植物进化历史中的系统发生过程等等,感兴趣的读者可以去搜索一下文献,某些知识分享网站上也有一些非常专业的回答。
下一篇再见!
参考文献:
蔡水花, 马文宝, 周晓, 等. 野生宝兴杜鹃重瓣花的发现及其形成机制探讨[J]. 四川林业科技, 2022, 43(01): 121-126.
付彦荣. 中国北方野生莲(Nelumbo nucifera)种质多样性研究[D]. 北京林业大学, 2021.
刘引. 月季种质资源评价及部分观赏性状QTL定位研究[D]. 河南农业大学, 2021.
彭洁, 刘引, 武荣花, 等. 重瓣花及其分子机制的研究进展[J]. 中国农学通报, 2023, 39(19): 65-72.
戚雨岑, 杨淑敏, 苏琳, 等. RcAGL61基因调控雄蕊和花瓣之间转变影响月季花瓣数量[J]. 西北植物学报, 2024, 44(06): 904-911.
孙明伟, 邵小斌, 赵统利, 等. 东方百合花器官数量变异分析[J]. 福建农业学报, 2018, 33(04): 373-380.
王宏哲, 张睿, 程劼, 等. 花基本结构的多样性及其分子机制[J]. 中国科学: 生命科学, 2019,49(04): 292-300.
王其超, 陈耀东, 辛春德, 等. 黑龙江野生荷花资源考察[J]. 中国园林, 1997(04): 39-41.
严寒松. 中国莲高密度遗传图谱的构建及花瓣数控制基因的定位[D]. 福建农林大学, 2019.
赵印泉, 刘青林. 重瓣花的形成机理及遗传特性研究进展[J]. 西北植物学报, 2009, 29(04): 832-841.
Annick D, OlivierR, Marion M, et al. Tinkering with the C-function: a molecular frame for theselection of double flowers in cultivated roses [J]. PloS one, 2010, 5(2): e9288.
De Craene LR. Meristicchanges in flowering plants: How flowers play with numbers[J]. Flora, 2016, 22-37.
Endress PK. TheFlowers in Extant Basal Angiosperms and Inferences on Ancestral Flowers[J]. InternationalJournal of Plant Sciences, 2001, 162(5): 1111-1140.
Endress PK,Doyle JA. Floral phyllotaxis in basal angiosperms: development andevolution[J]. Current opinion in plant biology, 2007, 10(1): 52-7.
Fan ML, Li XL,Zhang J, et al. Floral organ transcriptome in Camellia sasanqua providedinsight into stamen petaloid[J]. BMC Plant Biology, 2022, 22(1): 474-474.
Nan M, Wen C, TiangangF, et al. Low temperature-induced DNA hypermethylation attenuates expression ofRhAG, an AGAMOUS homolog, and increases petal number in rose (Rosa hybrida)[J]. BMC plant biology, 2015, 15(1): 237.
Reinhardt D,Gola EM. Law and order in plants - the origin and functional relevance ofphyllotaxis[J]. Trends in plant science, 2022, 27(10): 1017-1032.
Rümpler F, Theissen G. Reconstructing the ancestral flower of extantangiosperms: the 'war of the whorls' is heating up[J]. Journal of experimentalbotany, 2019, 70(10): 2615-2622.
Smyth DR. Evolutionand genetic control of the floral ground plan[J]. The New phytologist, 2018, 220(1):70-86.
Wang PP, Liao H, Zhang WG, et al. Flexibility inthe structure of spiral flowers and its underlying mechanisms[J]. NaturePlants, 2016, 2(1): 15188.